
J.
Phytiol.
(1976),
261,
pp.
387-422
387
With
2
plates
and
13
text-figure.
Printed
in
Great
Britain
POLYNEURONAL
INNERVATION
OF
SKELETAL
MUSCLE
IN
NEW-BORN
RATS
AND
ITS
ELIMINATION
DURING
MATURATION
BY
M.
C.
BROWN,*
J.
K.
S.
JANSEN
AND
D.
VAN
ESSENt
From
the
Inrtitute
of
Physiology,
Univer8ity
of
0810,
Karl
Johan8
Gate
47,
0810
1,
Norway
(Received
27
February
1976)
SUMMARY
1.
The
events
taking
place
during
the
elimination
of
polyneuronal
inner-
vation
in
the
soleus
muscle
of
new-born
rats
have
been
studied
using
a
combination
of
electrophysiological
and
anatomical
techniques.
2.
Each
immature
muscle
fibre
is
supplied
by
two
or
more
motor
axons
which
converge
on
to
a
single
end-plate.
There
was
no
sign
of
electrical
coupling
between
muscle
fibres
receiving
multiple
synaptic
inputs.
By
the
end
of
the
second
week
after
birth
virtually
all
muscle
fibres
are
inner-
vated
by
only
a
single
motor
axon.
3.
The
average
tension
produced
by
individual
motor
units,
measured
in
terms
of
the
percentage
of
the
total
muscle
twitch
tension,
declined
dramatically
during
the
first
2
weeks
after
birth.
During
this
period
there
was
no
significant
change
in
the
number
of
motor
neurones
innervating
the
soleus
muscle.
Thus,
the
disappearance
of
polyneuronal
innervation
reflects
a
decrease
in
the
number
of
peripheral
synapses
made
by
each
motor
neurone.
4.
The
decline
in
motor
unit
size
was
delayed,
but
not
ultimately
pre-
vented,
by
the
early
surgical
removal
of
all
but
a
few
motor
axons
to
the
soleus
muscle.
This
procedure
also
caused
a
delay
in
the
removal
of
poly-
neuronal
innervation
involving
the
remaining
motor
units.
5.
Following
a
crush
of
the
soleus
nerve
in
neonatal
animals,
regenera-
ting
axons
usually
returned
to
the
original
end-plates.
Polyneuronal
inner-
vation
was
extensive
at
early
stages
of
re-innervation
and
it
disappeared
during
the
second
week
after
birth
just
as
in
normal
muscles.
6.
Cross-innervation
of
neonatal
muscles
by
an
implanted
foreign
nerve
caused
a
rapid
disappearance
of
cholinesterase
at
denervated
original
*
Present
address:
University
Laboratory
of
Physiology,
Parks
Road,
Oxford.
t
Present
address:
Department
of
Anatomy,
University
College
London,
Gower
Street,
London
WC
1.

388
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
end-plates
and
in
most
fibres
prevented
re-innervation
by
the
original
nerve.
In
the
small
proportion
of
fibres
that
did
become
innervated
through
both
the
foreign
and
original
nerves
the
end-plates
were
more
than
1
mm
apart,
and
both
foreign
and
original
nerve
end-plates
could
persist
indefinitely.
7.
Many
cross-innervated
fibres
received
multiple
inputs
through
the
foreign
nerve.
Some
foreign
end-plates
were
separated
by
distances
ranging
up
to
1
mm.
Polyneuronal
innervation
through
the
foreign
nerve
was
completely
eliminated
during
maturation
but
over
a
slightly
longer
period
than
in
normal
muscles.
Apparently
the
elimination
process
can
act
over
a
distance
up
to
but
not
much
more
than
1
mm.
8.
These
observations
suggest
that
there
are
several
factors
influencing
the
elimination
of
redundant
inputs
in
immature
muscles.
Individual
motor
neurones
appear
to
have
an
inherent
tendency
to
withdraw
the
majority
of
their
original
complement
of
peripheral
terminals.
The
deter-
mination
of
which
particular
synapses
are
to
survive,
however,
seems
to
be
made
in
the
periphery
by
a
selection
among
all
the
synapses
that
inner-
vate
a
limited
region
of
each
muscle
fibre.
There
may
be
a
competitive
interaction
among
synapses
in
which
those
belonging
to
smaller
motor
units
are
less
likely
to
be
eliminated,
thereby
leading
to
a
relatively
uni-
form
size
of
the
motor
units
in
the
soleus.
INTRODUCTION
Mature
mammalian
skeletal
muscle
is
innervated
in
a
remarkably
simple
way
by
its
motor
axons,
in
the
sense
that
each
muscle
fibre
is
supplied
by
only
a
single
axon
terminal
situated
approximately
midway
along
the
length
of
the
fibre.
It
thus
came
as
a
surprise
when
Redfern
(1970)
demon-
strated
that
the
end-plate
potentials
(e.p.p.s)
of
muscle
fibres
from
new-
born
rats
consisted
of
several
discrete
components,
indicating
that
each
muscle
fibre
initially
receives
an
input
from
several
motor
axons.
During
the
first
few
weeks
after
birth
multiple
innervation
disappears
and
the
adult
pattern
of
single
innervation
of
each
muscle
fibre
is
achieved.
These
observations
have
been
confirmed
and
extended
by
Bennett
&
Pettigrew
(1974a);
Bagust,
Lewis
&
Westerman
(1973)
have
shown
that
a
similar
phenomenon
occurs
in
kitten
muscles.
The
accessibility
of
muscles
even
in
neonatal
animals
to
experimental
procedures
such
as
micro-electrode
recording,
partial
denervation,
re-
innervation,
and
cross-innervation
has
made
it
possible
to
ask
a
variety
of
questions
concerning
the
process
of
synapse
elimination.
For
example,
where
are
the
synaptic
terminals
located?
What
factors
determine
the
time
at
which
synapses
disappear:
is
it
related
specifically
to
the
age
of
the
muscle,
the
motor
neurone,
or
the
synapse?
Does
synapse
elimination

ELIMINATION
OF
SYNAPSES
involve
only
terminals
situated
very
close
to
one
another,
or
can
the
inter-
action
take
place
between
spatially
separated
terminals?
Do
immature
fibres
show
a
preference
for
retaining
innervation
from
their
original
nerve
supply
rather
than
from
a
foreign
nerve?
In
addition
to
providing
answers
to
some
of
these
and
related
questions,
our
results
give
clues
to
the
control
mechanisms
involved
in
producing
the
end
result
of
one
and
only
one
synapse
per
muscle
fibre.
Some
of
the
observations
have
been
presented
in
preliminary
communications
(Brown,
Jansen
&
Van
Essen
1975,
1976).
METHODS
Phy&iological
recordings.
Experiments
were
made
on
the
soleus
and
diaphragm
muscles
of
rats
of
different
ages,
starting
as
early
as
the
first
day
after
birth.
We
refer
to
muscles
from
animals
under
2
weeks
of
age
as
immature,
because
up
until
that
time
they
retain
a
pattern
of
innervation
clearly
different
from
that
seen
in
the
adult.
For
physiological
recordings
the
muscle
and
its
motor
nerve
supply
were
isolated,
pinned
out
in
a
small
chamber
coated
with
transparent
Sylgard
resin,
and
perfused
at
room
temperature
with
a
well-oxygenated
Ringer
solution
of
the
fol-
lowing
composition
(in
mM):
NaCl,
137;
KCl,
5;
CaCl1,
10;
MgCl2,
1;
glucose,
11;
Tris
Cl,
10;
buffered
to
pH
7-4.
The
high
Ca
in
the
solution
improved
the
stability
and
quality
of
micro-electrode
penetrations.
Intracellular
recordings
were
made
using
glass
micro-electrodes
filled
with
4
M
K-acetate
and
having
resistances
of
20-60
Mil.
Using
dark-field
illumination
it
was
possible
to
resolve
individual
muscle
fibres
even
in
the
small
immature
muscles.
Nerves
were
stimulated
through
glass-
tipped
suction
electrodes.
In
many
experiments
the
muscle
was
curarized
by
adding
just
enough
D-tubocurarine
(0-5-2
jug/ml.)
to
block
nerve-evoked
contractions.
In
experiments
involving
iontophoresis
of
acetylcholine
(ACh)
the
ACh
was
delivered
from
a
micropipette
filled
with
1
M
ACh
having
a
resistance
of
200-400
MKI.
A
backing
current
of
1-3
nA
was
used
to
minimize
the
desensitization
of
ACh
receptors.
Measurement
of
motor
unit
tension.
In
one
series
of
experiments
we
counted
the
total
number
of
motor
axons
supplying
the
soleus
muscle
and
the
tension
generated
by
individual
motor
units.
The
nerve
supply
to
the
soleus
muscle
was
dissected
free
all
the
way
back
to
spinal
roots
L5
and
L4.
The
preparation
was
then
transferred
to
a
chamber
having
two
compartments
separated
by
a
thin
plastic
partition.
The
soleus
nerve
was
placed
through
a
slot
in
the
partition,
and
the
proximal
tendon
of
the
muscle
was
pinned
to
the
bottom
of
the
chamber
close
to
the
partition.
The
distal
tendon
was
connected
by
fine
surgical
thread
to
a
sensitive
strain
gauge.
Activity
in
the
soleus
nerve
was
monitored
by
recording
differentially
from
the
two
compart-
ments,
which
were
perfused
independently.
Each
ventral
root
was
split
into
several
filaments
that
were
stimulated
individually.
The
splitting
was
continued
until
each
filament
contained
at
most
four,
and
usually
only
one
or
two
soleus
motor
axons.
We
occasionally
saw
action
potentials
in
the
soleus
nerve
not
followed
by
measurable
tension
changes
in
the
muscle,
but
these
were
discounted
because
they
were
pre-
sumably
either
from
y
motor
axons
or
from
gastrocnemius
motor
axons
extending
unusually
far
into
the
soleus
nerve.
For
direct
stimulation
current
pulses
were
passed
between
coarse
electrodes
placed
on
opposite
sides
of
the
muscle.
In
the
course
of
most
experiments
there
was
an
unexplained
slow
and
parallel
decline
of
10-30%
in
the
tensions
produced
by
direct
muscle
stimulation
and
by
maximal
stimulation
of
the
nerve.
When
this
occurred
the
individual
motor
unit
tensions
were
expressed
as
a
percentage
of
the
original
total.
The
decline
in
muscle
tension,
389

390
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
whatever
its
cause,
would
have
only
a
slight
effect
on
our
estimates
of
average
motor
unit
size.
It
might,
however,
seriously
affect
our
measurements
of
the
total
range
in
motor
unit
size
if
the
over-all
tension
decline
involved
large
losses
by
only
a
few
motor
units.
This
seems
unlikely,
though,
as
small
motor
units
were
encountered
even
in
experiments
where
little
or
no
tension
loss
was
seen
and
also
at
early
stages
of
the
other
experiments,
before
the
tension
decline
had
set
in.
Hieological
techniques.
End-plate
cholinesterase
was
stained
using
the
procedure
of
Karnovsky
(1964).
Muscles
were
prefixed
for
10-15
min
in
a
solution
containing
2-
5
%
glutaraldehyde,
2
%
paraformaldehyde,
and
0-1
IM
cacodylate
buffer
at
pH
7-5.
They
were
then
incubated
in
the
staining
solution
at
room
temperature
for
30
min,
fixed
in
the
same
fixative
solution
for
1-24
hr,
and
stored
in
H20.
The
distri-
bution
of
end-plates
throughout
the
muscle
could
be
seen
easily
in
whole
mount
preparations
viewed
under
a
dissecting
microscope.
In
order
to
count
the
number
of
end-plates
on
individual
fibres
we
found
it
necessary
to
tease
out
single
fibres
over
the
entire
region
where
cholinesterase
was
visible
in
the
whole
mount.
Fibres
were
dissected
free
with
the
aid
of
electrolytically
sharpened
tungsten
needles
and
fine
dissecting
forceps.
Each
fibre
was
placed
in
Aquamount
under
a
cover-slip
and
viewed
at
high
power
in
order
to
resolve
individual
end-plates
and
to
ensure
that
only
a
single
fibre
had
been
isolated.
Nerve
terminals
and
their
preterminal
axons
were
stained
using
the
zinc
iodide-
osmium
technique
(Akert
&
Sandri
1968).
Muscles
were
incubated
16-18
hr
in
the
staining
solution
and
washed
in
H20
for
at
least
2
hr.
Frozen
sections
of
30
jam
thick-
ness
were
cut
parallel
to
the
length
of
the
muscle
and
mounted
in
Aquamount.
The
staining
of
nerve
terminals
was
more
consistent
in
the
diaphragm
than
in
the
soleus
muscle,
presumably
because
the
diaphragm
is
a
much
thinner
sheet
of
muscle.
Surgical
procedures.
Operations
were
carried
out
under
ether
anaesthesia,
usually
on
the
first
or
second
day
after
birth.
The
soleus
muscle
was
denervated
by
crushing
the
nerve
next
to
its
entry
into
the
muscle
and
cross-innervated
by
placing
the
superficial
branch
of
the
fibular
nerve
on
to
the
proximal
surface
of
the
muscles.
The
effectiveness
of
the
nerve
crush
in
completely
interrupting
the
original
nerve
supply
was
demonstrated
by
showing
that
at
early
stages
(up
to
a
week
after
the
operation)
most
muscle
fibres
were
still
denervated,
and
that
those
which
were
in-
nervated
had
end-plate
potentials
with
abnormally
long
latencies
owing
to
the
slower
conduction
velocity
of
regenerated
axons.
Furthermore,
cross-innervation,
which
did
not
occur
when
the
original
nerve
was
left
intact,
took
place
just
as
effectively
after
a
nerve
crush
as
after
a
cut.
Nerve
crushes
were
preferred
for
most
experiments
because
re-innervation
took
place
more
quickly
and
reliably
than
after
a
nerve
cut.
In
one
series
of
experiments
the
soleus
muscle
was
partially
denervated
between
day
3
and
day
7
after
birth
by
cutting
ventral
root
L5,
which
contains
all
but
a
few
of
the
motor
axons
supplying
the
muscle.
The
root
was
cut
either
within
or
just
outside
the
spinal
column.
Most
animals
survived
the
operation
but
since
it
was
not
possible
to
recognize
the
nerve
roots
individually
during
the
operation
L5
was
successfully
cut
and
L4
left
undamaged
in
only
nine
animals
out
of
more
than
fifty
upon
which
we
operated.
RESULTS
Normal
neonatal
muscle
The
compound
end-plate
potential
In
muscles
taken
from
animals
less
than
10
days
old
virtually
all
muscle
fibres
receive
synaptic
inputs
from
several
motor
axons.
Text-
fig.
1
shows,
for
example,
an
intracellular
recording
from
a
fibre
in
a

ELIMINATION
OF
S
YNAPSES
5-day-old
soleus
muscle
paralysed
with
curare.
By
grading
the
strength
of
the
stimulus
to
the
nerve
it
was
possible
to
demonstrate
three
distinct
components
of
the
e.p.p.
The
nerve
was
stimulated
twice
in
each
of
the
six
superimposed
traces
in
the
Text-figure.
The
first
stimulus
was
varied
in
strength,
progressively
recruiting
one,
two
and
then
three
discrete
inputs;
the
second
stimulus
was
kept
supramaximal
to
indicate
the
degree
of
variability
in
maximal
e.p.p.
amplitude
from
one
trial
to
the
next.
The
Aft~~~~~~~~~~
Graded
Maximal
4mV
stimuli
stimuli
20
msec
Text-fig.
1.
The
compound
e.p.p.
Intracellular
recording
(a.c.)
from
a
soleus
muscle
fibre
of
a
5-day-old
rat.
The
soleus
nerve
was
stimulated
twice
during
each
of
six
superimposed sweeps.
The
first
shock
was
graded
in
strength,
while
the
second
was
kept
supramaximal.
Each
of
the
three
different
e.p.p.
components
was
recruited
at
sharply
defined
threshold
levels.
The
muscle
was
paralyzed
with
D-tubucurarine,
1
jcglml.
different
components
of
the
e.p.p.
invariably
had
very
similar
rise
times
which
were
as
short
as
2
msec
when
the
recording
electrode
was
situated
focally.
Often
all
of
the
components
had
a
similar
amplitude,
as
in
Text-
fig.
1,
but
occasionally
their
sizes
differed
by
as
much
as
a
factor
of
ten.
These
observations
are
in
basic
agreement
with
those
of
Redfern
(1970)
and
of
Bennett
&
Pettigrew
(1974a),
except
that
Bennett
&
Pettigrew
occa-
sionally
saw
e.p.p.
components
having
different
time
courses
in
muscles
examined
before
the
end
of
gestation.
Absence
of
electrical
coupling
between
muscle
fibres
One
simple
explanation
for
the
presence
of
several
components
in
the
e.p.p.
is
that
immature
muscle
fibres
might
be
electrically
coupled
to
one
another,
a
phenomenon
which
has
been
demonstrated
in
regenerating
salamander
muscle
(Dennis,
1975)
and
in
developing
amphibian
muscle
(Blackshaw
&
Warner,
1976).
In
the
rat
the
coupling
would
have
to
be
strong
in
order
to
account
for
the
equality
in
rise
times
and
similarity
in
amplitudes
of
the
different
components
of
the
e.p.p.
We
were
not,
how-
ever,
able
to
detect
any
signs
of
coupling
between
immature
muscle
fibres
in
extensive
surveys
of
several
muscles
examined
in
the
first
2
weeks
391

392
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
after
birth.
When
two
adjacent
muscle
fibres
were
impaled
with
separate
micro-electrodes,
current
injected
into
one
fibre
never
caused
a
measur-
able
potential
change
in
the
neighbouring
fibre,
even
when
the
stimulus
was
sufficient
to
set
up
an
action
potential
in
the
first
fibre.
This
result
rules
out
any
widespread
electrical
coupling
in
the
immature
soleus
muscle,
but
a
more
restricted
alternative,
that
of
coupling
within
small
groups
of
muscle
fibres,
remained
a
possibility
if
one
supposed
that
most
of
the
fibres
within
a
group
were
too
small
to
be
clearly
resolved
in
the
dissecting
microscope.
We
therefore
carried
out
other
experiments
to
elucidate
the
pattern
of
innervation
of
neonatal
muscle
fibres.
A
10
mV/
20
msec
Text-fig.
2.
The
effect
of
prostigmine
on
the
compound
e.p.p.
A,
rapidly
rising
and
decaying
e.p.p.s
(single
component
on
left,
double
on
right)
in
the
absence
of
prostigmine
in
a
10-day
muscle
paralysed
with
curare,
1
jug/ml.
B,
slowly
rising
and
decaying
e.p.p.
components
recorded
from
the
same
fibre
several
minutes
after
applying
prostigmine,
2
x
10-6
gfml.
Multiple
innervation
of
single
end-plate
sites
Staining
of
a
neonatal
soleus
muscle
for
cholinesterase
reveals
that
the
end-plates
are
distributed
along
a
narrow
band
that
lies
roughly
midway
along
the
length
of
the
muscle,
just
as
is
found
in
adult
muscles.
In
whole
mounts
and
in
sectioned
material,
it
is
difficult
to
tell
whether
adjacent
spots
of
cholinesterase
are
on
the
same
or
on
neighbouring
fibres.
Lubinska
&
Zelena
(1966)
found,
however,
only
a
single
spot
of
cholinesterase
on
single
muscle
fibres
teased
from
the
new-born
rat
diaphragm.
We
have
confirmed
this
point
for
the
immature
soleus
muscle
(PI.
1
B).

ELIMINATION
OF
SYNAPSES
We
tested
for
the
possibility
that
some
synapses
might
lack
end-plate
cholinesterase
altogether
by
examining
the
effects
of
prostigmine
on
the
time
course
of
e.p.p.s
in
partially
curarized
muscles.
In
the
experiment
illustrated
in
Text-fig.
2
the
application
of
prostigmine
(2
,Ug/m1.)
to
the
bathing
solution
more
than
doubled
the
rise
times
and
amplitudes
of
both
the
low-threshold
e.p.p.
component
elicited
by
the
first
stimulus
and
the
maximal
e.p.p.
evoked
by
the
second
stimulus.
Prostigmine
had
a
similar
effect
on
all
components
of the
e.p.p.
in
every
muscle
fibre
tested,
indi-
cating
that
there
is
cholinesterase
associated
with
all
of
the
synapses
present
in
immature
muscles.
Since
only
one
spot
on
each
fibre
stains
for
cholinesterase,
it
is
likely
that
all
its
terminals
are
situated
at
this
one
end-plate.
Further
evidence
for
convergent
innervation
from
several
motor
axons
came
from
examining
muscles
whose
nerve
terminals
had
been
stained
using
the
zinc
iodide-osmium
technique.
In
well-stained
sections
from
immature
muscles
(both
the
soleus
and
the
diaphragm)
it
was
usually
possible
to
see
two
or
more
axons
leading
into
each
end-plate
(PI.
1A,
arrows).
In
older
muscles
examined
20
or
more
days
after
birth,
when
physiological
signs
of
multiple
innervation
have
virtually
disappeared,
there
was
only
one
axon
supplying
each
end-plate.
These
observations,
which
are
in
agreement
with
those
made
by
Bennett
&
Pettigrew
(1974a)
on
silver-stained
muscles,
provide
strong
support
for
the
idea
that
each
end-plate
initially
receives
an
input
from
several
motor
axons.
Strictly
speaking,
though,
the
anatomical
evidence
is
not
by
itself
conclusive
because
we
were
not
able
to
trace
the
preterminal
axons
to
each
fibre
far
enough
back
towards
the
main
nerve
to
be
certain
that
they
were
no
branches
of
the
same
parent
axon.
Our
final
piece
of
evidence
concerning
the
distribution
of
synapses
in
immature
muscles
comes
from
experiments
in
which
localized
application
of
ACh
from
a
micropipette
was
used
to
set
an
upper
limit
to
the
separation
between
synapses
on
each
fibre.
The
principle
of
the
experiment
was
to
see
whether
desensitization
of
the
ACh
receptors
over
a
small
region
of
a
muscle
fibre
had
an
equal
effect
on
all
components
of
the
e.p.p.
The
re-
cording
micro-electrode
was
first
inserted
into
a
muscle
fibre
near
the
synaptic
region
in
a
curarized
muscle
and
multiple
components
of
the
e.p.p.
were
demonstrated
by
the
paired
stimulation
technique
described
above
(Text-fig.
3A).
The
ACh
micropipette
was
then
moved
in
small
steps
along
the
fibre
until
a
spot
was
found
having
a
moderately
high
sensitivity
to
ACh
(>
10
mV/nC
in
the
presence
of
curare,
1-2
x
10-6
g/ml.).
At
this
point
a
steady
positive
current
of
1-5
nA
through
the
ACh
pipette
invariably
caused
a
large
reduction
or
even
a
complete
abolition
of
all
components
of
the
e.p.p.
(Text-fig.
3
B).
Such
a
parallel
reduction
393

394
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
of
all
components
of
the
e.p.p.
was
seen
in
every
experiment.
The
ACh
application
also
caused
a
steady
conductance
change
and
membrane
depolarization
(Text-fig.
3,
lower
traces),
both
of
which
would
contribute
indirectly
to
the
reduction
in
e.p.p.
amplitude.
The
membrane
potential
always
remained
well
below
the
reversal
potential
for
the
e.p.p.,
however,
even
during
ACh
applications
that
completely
abolished
the
e.p.p.;
moreover,
considerable
reductions
in
e.p.p.
amplitude
could
be
obtained
by
ACh
applications
that
depolarized
the
membrane
by
only
a
few
milli-
volts.
It
is
clear,
therefore,
that
the
parallel
effects
of
ACh
application
on
all
components
of
the
e.p.p.
must
have
resulted
primarily
from
a
direct
desensitization
of
ACh
receptors
at
the
end-plate.
A
B
C
0
.
F
~~~~~~~~~~~
~~~5
mV
Eli
-40
20
msec
Text-fig.
3.
Desensitization
of
end-plate
receptors
by
ACh
iontophoresis.
A,
compound
e.p.p.
from
a
10-day-old
curarized
soleus
muscle
fibre.
High
gain,
a.c.
records
above;
low
gain,
d.c.
records
below.
B,
all
components
of
the
e.p.p.
were
abolished
during
steady
iontophoretic
application
of
ACh
to
the
end-plate
region.
C,
both
e.p.p.
components
recovered
completely
several
seconds
after
the
cessation
of
ACh
ionophoresis.
Preparation
curarized,
1
ug/ml.
Careful
positioning
of
the
ACh
pipette
was
necessary
in
order to
obtain
this
desensitizing
effect.
Movements
of
the
pipette
by
30-40
/Um
in
either
direction
along
the
length
of
the
fibre
or
over
to
adjacent
fibres
greatly
reduced
or
abolished
the
effect.
Although
the
spatial
resolution
provided
by
the
technique
as
we
used
it
was
somewhat
coarser
than
the
dimensions
of
an
end-plate
or
the
diameter
of
an
immature
muscle
fibre,
it
is
neverthe-
less
sufficient
to
demonstrate
that
all
of
the
synapses
on
an
immature
muscle
fibre
are
situated
within
about
50
gsm
of
one
another.
The
elimination
of
polyneuronal
innervation
The
percentage
of
muscle
fibres
receiving
more
than
one
synaptic
input
declined
rapidly
during
the
second
week
after
birth.
Text-fig.
4
shows
the
results
from
twenty-six
muscles
examined
between
the
second
and
nine-
teenth
day
after
birth.
At
least
twenty
fibres
from
each
muscle
were
tested
for
multiple
inputs
in
the
manner
illustrated
in
Text-fig..
1.
During
the
5
day
period
from
day
10
to
15
the
percentage
of
fibres
having
multiple

ELIMINATION
OF
SYNAPSES
inputs
dropped
from
91
%
(forty
of
forty-four)
fibres
to
25
%
(one
of
forty
fibres).
The
incidence
of
fibres
having
three
or
more
components
was
clearly
higher
during
the
first
week
after
birth
than
in
the
second
week,
but
we
did
not
examine
this
point
systematically
because
the
fluctuations
in
maximal
e.p.p.
amplitude
(which
were
usually
greater
than
that
shown
in
Text-fig.
1)
often
made
it
difficult
to
determine
exactly
how
many
components
were
present.
The
elimination
of
multiple
synaptic
inputs
100
_
-@
--
*@@
°,
50_
.
0~~~~~~~
25_\
>
75
0
5
10
15
20
Age
(days)
Texrt-fig.
4.
Percentage
of
soleus
muscle
fibres
innervated
by
more
than
one
axron
at
different
ages,
determined
from
intracellular
recordings
in
curar-
ized
muscles.
At
least
twenty
fibres
were
examined
in
each
muscle.
Treor
more
e.p.p.
components
were
seen
in
many
fibres,
especially
at
early
ages,
suggesting
that
the
loss
of
extra
synapses
may
be
well
under
way
before
day
10.
Continuous
line
drawn
by
eye
to
fit
the
observations.
thus
appears
to
be
an
ongoing
process
that
starts
sometime
during
the
first
week
after
birth
and
continues
until
the
end
of
the
second
week
(see
also
Bennet
&;
Pettigrew,
1974a
and
Texrt-fig.
6
below).
After
this
time
we
found
a
low
level
of
multiple
innervation
remaining
for
a
few
days,
but
even
this
must
eventually
disappear
(see
Jansen
&;
Van
Essen,
1975).
During
the
period
when
multiple
synaptic
inputs
were
being
eliminated
there
was
no
sign
that
significant
numbers
of
muscle
fibres
became
tran-
siently
denervated.
Once
the
recording
micro-electrode
was
placed
in
a
region
where
focal
e.p.p.s.
could
be
recorded
it
was
usually
possible
to
record
long
sequences
of
twenty
or
more
fibres
all
having
normal
e.p.p.s
395

396
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
with
one
or
more
components.
In
the
few
cases
where
one
or
more
fibres
showing
no
response
to
nerve
stimulation
were
encountered
it
was
possible
to
attribute
the
change
either
to
a
sudden
shift
in
the
location
of
the
end-
plates
or
to
a
widespread
block
in
neuromuscular
transmission.
This
result
suggests
that
if
there
is
a
period
of
transient
denervation
during
the
matur-
ation
of
the
muscle,
it
either
involves
very
few
fibres
or
else
lasts
for
a
very
short
time
before
the
fibre
disappears
or
is
re-innervated.
Number
and
size
of
motor
units
The
removal
of
hyperinnervation
during
maturation
could
be
associated
with
the
complete
loss
of
whole
motor
neurones,
by
a
reduction
in
the
number
of
peripheral
terminations
made
by
each
motor
neurone,
or
by
some
combination
ofthe
two.
Degeneration
of
presumptive
motor
neurones
is
a
well
known
phenomenon
during
early
stages
of
development
(Hughes,
1968;
Landmesser
&
Pilar,
1974),
and
Bennett
&
Pettigrew
(1974a)
suggested
that
this
might
explain
the
elimination
of
synapses
during
the
early
post-natal
period.
The
question
was
settled
by
determining
the
size
and
the
total
number
of
motor
units
supplying
the
soleus
muscle
at
differ-
ent
ages.
On
account
of
the
rapid
growth
of
the
muscle
during
matura-
tion,
motor
unit
size
is
defined
operationally
as
the
percentage
of
the
total
muscle
twitch
tension
produced
by
that
unit.
Since
fibre
diameters
are
relatively
uniform
in
the
soleus
the
tension
measurements
should
provide
a
reasonable
indication
of
the
percentage
of
muscle
fibres
innervated
by
each
motor
unit.
In
order
to
count
the
total
number
of
soleus
motor
units,
the
muscle
was
dissected
free
along
with
its
nerve
supply
all
the
way
back
to
the
spinal
cord
and
mounted
in
a
chamber
that
allowed
impulse
activity
in
the
soleus
nerve
and
tension
in
the
muscle
to
be
recorded
while
small
filaments
of
the
ventral
roots
were
stimulated
electrically
(see
Methods).
We
found
that
the
total
number
of
motor
units
remained
virtually
constant
throughout
the
period
when
polyneuronal
innervation
was
being
eliminated.
This
number
was
between
twenty-one
and
twenty-five
for
the
seven
muscles
between
3
and
42
days
of
age
for
which
we
obtained
complete
counts.
There
was
no
significant
difference
in
the
number
of
motor
units
for
the
three
muscles
examined
before
day
10
(mean
22-7
units)
and
the
four
muscles
examined
after
that
time
(mean
23x5
units).
A
few
motor
units
may
have
been
missed
in
some
of
the
preparations;
in
fact,
our
value
for
mature
muscle
is
slightly
lower
than
the
estimates
of
others
based
on
in
vivo
tension
measurements
(28-30
units;
Close,
1967)
or
fibre
counts
in
deafferented
motor
nerves
(thirty-two
a
motor
axons;
Gutman
&
Hanz-
llkova',
1966).
Nevertheless,
it
is
unlikely
that
we
missed
a
much
greater
number
of
units
in
younger
than
in
older
rats
and
we
therefore
conclude

ELIMINATION
OF
S
YNAPSES
that
the
number
of
motor
units
to
the
soleus
stays
relatively
constant
after
birth.
In
contrast
to
the
constancy
of
the
number
of
motor
units,
there
were
dramatic
changes
in
motor
unit
size
shortly
after
birth.
Text-fig.
5,
for
example,
shows
a
single
motor
unit
from
an
immature
muscle
that
gener-
ated
a
tension
greater
than
one
fifth
of
the
total
muscle
tension.
Text-fig.
5B
is
a
recording
from
the
soleus
nerve
at
a
fast
sweep
speed
to
demon-
strate
that
the
stimulus
activated
only
a
single
soleus
motor
axon.
Text-
fig.
5A
shows,
at
a
much
slower
sweep
speed,
the
tension
generated
by
the
Maximal
tension
g
wt..
I
_LZ
Single
motor
unit
tension
2001
PV
|
Nerve
500
msec
50
msec
Text-fig.
5.
Measurement
of
motor
unit
size
in
an
immature
muscle.
A,
shows
the
tension
generated
in
the
soleus
muscle
of
a
3-day-old
rat
by
stimulation
of
a
single
ventral
root
filament
and,
for
comparison,
the
ten-
sion
produced
by
maximal
stimulation
of
the
whole
nerve.
B,
shows,
at
a
fast
sweep
speed,
the
unitary
action
potential
recorded
en
pa88age
from
the
soleus
nerve
(see
Methods)
after
stimulation
of
the
ventral
root
filament.
Note
the
very
slow
time
course
of
the
contraction,
which
is
characteristic
of
immature
muscle.
single
motor
unit
and,
for
comparison,
the
total
muscle
tension
produced
by
maximal
stimulation
of
the
nerve.
(Direct
electrical
stimulation
of
the
muscle
also
produced
the
same
maximal
tension.)
Our
observations
on
motor
unit
tensions
are
summarized
in
Text-fig.
6,
which
gives
the
relative
tension
of
the
motor
units
in
ten
different
muscles
as
a
function
of
the
age
of
the
animal.
The
tensions
are
displayed
as
per-
centages
of
the
total
twitch
tension
evoked
by
direct
stimulation
of
the
muscle.
Each
vertical
line
represents
the
observations
in
one
animal,
and
the
mean
values
(filled
circles)
as
well
as
the
values
for
individual
units
(horizontal
lines)
are
presented
for
each
muscle.
Tension
measurements
397

398
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
were
included
only
for
the
lowest
threshold
unit
in
each
ventral
root
filament
because
there
was,
as
expected,
non-linear
summation
of
tension
values
when
more
than
one
unit
was
stimulated.
The
open
circles
show
the
average
tension
that
would
be
expected
if
there
were
no
multiply
inner-
35
.
30
0
C
~25
0
20
20
0
CI
10
0
10
Is
20
4
Age
(days)
Text-fig.
6.
Size
and
number
of
soleus
motor
units
at
different
ages.
Each
vertical
line
represents
observations
on
one
animal.
The
ordinate
gives
the
size
of
motor
units
expressed
as
their
percentage
of
the
maxima
twitch
to
direct
stimulation.
Filled
circles
(X)
give
the
mean
size
of
motor
units
in
each
muscle;
horizontal
lines
indicate
the
individual
measurements
for
the
lowest-threshold
motor
unit
in
each
ventral
root
filament.
Open
circles
(0)
show
the
average
motor
unit
size
one
would
expect
in
the
absence
of
poly-
neuronal
innervation
[100
x
(total
number
of
units-l)].
In
three
muscles
(2,
5
and
17
days)
this
value
is
not
given
because
a
partial
nerve
block
in
the
region
of
the
ventral
roots
prevented
completion
of
the
motor
unit
count.
vated
fibres.
This
value,
obtained
simply
by
taking
the
inverse
of
the
total
number
of
motor
units,
was
between
4
and
5
%
for
the
seven
muscles
for
which
complete
motor
unit
counts
were
available.
Text-fig.
6
shows
that
motor
units
were
about
five
times
larger
at
early

ELIMINATION
OF
SYNAPSES
times
(mean
23
%
at
days
2
and
3)
than
at
later
times
(mean
5-1
%
at
days
15-18
and
4.4
%
at
day
42).
This
reduction
in
motor
unit
size
is
large
enough
to
account
entirely
for
the
disappearance
of
polyneuronal
innerva-
tion
in
immature
muscles.
In
fact,
the
average
degree
of
polyneuronal
innervation
estimated
from
the
motor
unit
measurements
(obtained
by
taking
the
ratio
of
the
filled
circle
to
the
open
circle
values
for
each
muscle),
is
about
five
at
days
2
and
3.
An
average
value
of
five
synapses
per
fibre
is
higher
than
either
we
or
Bennett
&
Pettigrew
(1974a)
obtained
by
count-
ing
the
number
of
e.p.p.
components
in
individual
fibres
but
the
dis-
crepancy
is
not
surprising
in
view
of
the
inherent
inaccuracies
in
each
type
of
estimate.
A
safe
conclusion
would
be
that
there
are
many
separate
synaptic
inputs
to
each
neonatal
muscle
fibre,
each
one
of
which
is
capable
of
activating
the
muscle
fibre
by
itself.
It
is
interesting
that
there
was
considerably
more
scatter
in
the
size
of
motor
units
throughout
the
period
of
maturation
than
in
adult
muscles.
The
total
range
was
about
a
factor
of
three
in
the
one
adult
muscle
we
examined
and
in
the
larger
sample
of
motor
units
obtained
by
Close
(1967),
whereas
the
range
was
about
tenfold
in
several
of
the
immature
muscles.
At
all
ages
there
were
some
motor
units
that
were
within
the
range
seen
in
the
adult.
In
the
15-
and
17-day-old
muscles
a
few
of the
motor
units
were
actually
smaller
than
any
seen
in
the
adult.
It
is
possible
that
the
tension
values
were
spuriously
low
owing
to
an
axonal
conduction
block
or
to
a
failure
in
synaptic
transmission.
This
explanation
seems
unlikely,
however,
because
the
maximal
indirect
and
direct
twitches
were
equal
and
in
the
same
muscles
there
were
still
other
motor
units
larger
than
those
seen
in
the
adult.
If
the
tension
measurements
provide
a
reasonably
accurate
measure
of
motor
unit
size
in
terms
of
numbers
of
muscle
fibres
innervated,
then
the
results
suggest
that
at
a
time
when
almost
all
polyneuronal
innervation
has
been
eliminated,
some
readjustments
in
motor
unit
sizes
remain
to
be
completed.
Modifying
the
time
course
of
synarpse
removal
The
effects
of
partial
denervation
If
one
could
remove
all
but
a
few
of
the
motor
axons
to
the
soleus
muscle
shortly
after
birth,
a
reduction
in
size
of
the
remaining
ones
would
have
to
take
place
at
the
expense
of
leaving
some
fibres
completely
de-
nervated.
A
priori
it
seemed
plausible
that
this
might
influence
or
even
reverse
the
normal
reduction
in
motor
unit
size.
The
experiment
was
performed
by
sectioning
the
lower
of
the
two
ventral
roots
supplying
the
soleus
muscle.
The
remaining
root
(L4)
usually
contains
at
least
one
and
sometimes
as
many
as
nine
soleus
motor
axons.
The
operation
was
carried
out
successfully
on
nine
animals.
They
were
examined
between
days
15
and
43
after
birth.
The
results
were
quite
striking:
partial
denervation,
on
399

400
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
the
one
hand,
delayed
the
normal
reduction
in
motor
unit
size
and
even
the
elimination
of
polyneuronal
innervation;
on
the
other
hand,
it
did
not
seem
to
prevent
the
eventual
shrinkage
of
motor
units
to
approximately
their
normal
adult
size.
Five
muscles
with
five
to
nine
remaining
motor
units
were
examined
between
days
15
and
17,
just
after
the
time
when
most
polyneuronal
innervation
normally
has
been
eliminated.
The
histogram
in
Text-fig.
7
l
l
'5
0
0
10
20
30
Motor
unit
sizes
Text-fig.
7.
Motor
unit
sizes
at
intermediate
times
after
partial
denerva-
tion.
Continuous
lines:
motor
unit
sizes,
determined
from
tension
measure-
ments
(as
in
Text-fig.
6),
of
twenty
motor
units
in
five
muscles
examined
15-17
days
after
birth
following
partial
denervation
on
day
3-5
(three
on
day
3,
one
each
on
days
4
and
5).
In
each
animal
there
were
between
four
and
nine
motor
units
in
the
remaining
ventral
root
(L4)
but
tension
meas-
urements
were
not
obtained
for
all
of
these.
Dashed
lines:
motor
unit
sizes
in
two
normal
soleus
muscles
aged
15
and
17
days
(from
Text-fig.
6).
shows
the
tensions
generated
by
twenty
of
the
motor
units
remaining
in
these
five
muscles.
The
sizes
of
seventeen
units
from
two
normal
muscles
of
comparable
age
are
shown
for
comparison.
The
mean
size
of
the
motor
units
from
the
partially
denervated
muscles
(20
%)
was
much
larger
than
that
of
the
normal
muscles
(5-6
%)
and
was,
in
fact,
very
close
to
the
mean
size
of
motor
units
at
the
time
of
the
initial
operation
(21
%;
results
from
days
3
and
5
in
Text-fig.
6).
The
abnormally
large
motor
units
did
not
appear
to
result
simply
from
hypertrophy
of
innervated
muscle
fibres
or
to
atrophy
of
the
denervated
ones.
The
muscles
were
not
noticeably
atrophic

ELIMINATION
OF
SYNAPSES
and
they
generated
a
normal
tension
to
nerve
stimulation.
In
addition,
cross-sections
of
two
of
the
muscles
showed
that
they
contained
close
to
the
normal
number
of
muscle
fibres
(2500-2900)
having
relatively
uniform
diameters.
Thus,
it
appears
that
the
motor
units
remaining
after
partial
denervation
innervated
a
much
larger
number
of
muscle
fibres
than
they
normally
would
have
at
this
age.
There
were
two
other
obvious
abnormalities
in
the
partially
denervated
muscles
examined
between
days
15
and
17.
The
first
was
that the
twitch
contractions
to
stimulation
of
single
motor
units
fatigued
very
rapidly,
even
during
stimulation
at
frequencies
as
low
as
0
5-1
Hz.
After
a
few
stimuli
the
twitch
tension
was
often
less
than
half
of
its
initial
value,
after
which
it
remained
stable.
In
normal
muscles
there
was
little
tension
decre-
ment
even
during
stimulation
at
rates
of
2-3
Hz.
The
rapid
fatigue
in
the
partially
denervated
muscles
was
not
related
to
an
impairment
in
the
con-
tractile
properties
of
the
muscle
as
there
was
no
fatigue
during
direct
muscle
stimulation.
A
likely
explanation
for
the
fatigue
is
that
transmission
at
many
synapses
was
only
marginally
above
threshold
and
was
readily
brought
below
threshold
by
repetitive
stimulation.
The
other
abnormality
in
this
group
of
partially
denervated
muscles
was
that
there
was
extensive
polyneuronal
innervation
remaining
2
weeks
after
the
operation.
We
recorded
e.p.p.s.
having
more
than
one
component
in
about
half
of
the
innervated
fibres
in
each
muscle
(range
22-65
%
in
five
muscles).
This
was
not
a
general
'systemic'
consequence
of the
opera-
tion,
because
in
every
experiment
we
found
that
polyneuronal
innerva-
tion
in
the
control
soleus
muscle
from
the
opposite
leg
had
been
reduced
to
its
normal
level
of
5
%
or
less.
The
situation
was
quite
different
in
muscles
examined
at
later
times
after
partial
denervation.
The
clearest
illustration
of
this
came
from
a
muscle
partially
denervated
four
days
after
birth
and
examined
on
day
33.
There
were
only
two
motor
axons
supplying
the
soleus
muscle
through
the
intact
ventral
root
and
the
muscle
itself
had
atrophied
considerably.
A
cross-section
of
the
muscle,
shown
in
P1.
2
B,
revealed
that
a
minority
of
the
muscle
fibres,
321
in
all,
had
escaped
atrophy
and
were
actually
larger
than
normal.
The
other
fibres
were
very
small
and
difficult
to
resolve
individually.
For
comparison,
P1.
2A
shows
the
relative
uniformity
of
fibre
diameters
in
a
normal
muscle
of
similar
age.
We
found
no
multiply
innervated
fibres
during
a
micro-electrode
survey
of
the
partially
dener-
vated
muscle
after
it
was
curarized.
Hence,
a
reasonable
estimate
of
motor
unit
sizes
can
be
obtained
by
assuming
that
the
very
small
fibres
in
the
muscle
were
all
denervated
and
then
simply
dividing
up
the
number
of
larger
fibres
in
accordance
with
the
relative
sizes
of
the
motor
unit
ten-
sions.
The
two
motor
units
in
this
muscle
contributed
40
and
60
%
to
the
401

402
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
total
muscle
tension,
suggesting
that
they
innervated
approximately
130
and
190
muscle
fibres,
respectively.
These
values
are
about
4
and
6
%
of
the
total
of
approximately
3000
fibres
found
in
normal
muscles
(Chiakulas
&
Pauly,
1965;
Frank,
Jansen,
Lomo
&
Westgaard,
1975).
A
similar
estimate
was
obtained
indirectly
by
comparing
the
absolute
values
of
the
motor
unit
tensions
obtained
from
normal
muscles
of
comparable
age.
The
total
twitch
tension
from
the
partially
denervated
muscle
was
about
15
%
of
that
from
normal
muscles
of
a
similar
age
(12
g
wt.
vs.
6'9-10
g
wt.
for
five
normal
4-6-week-old
muscles),
and
the
individual
motor
unit
tensions
were
roughly
6
and
9
%
of
the
normal
total.
Thus,
even
without
a
correction
for
the
effects
of
the
obvious
hypertrophy
of
innervated
muscle
fibres,
the
tension
measurements
support
the
suggestion
that
the
number
of
muscle
fibres
innervated
by
each
motor
unit
was
closer
to
the
mean
value
in
normal
adult
muscle,
than
to
the
mean
value
at
the
time
of
the
initial
operation
or
after
2
weeks
of
partial
denervation.
Owing
to
the
large
scatter
in
individual
motor
unit
sizes
in
immature
muscles,
however,
it
is
not
possible
to
conclude
that
these
particular
motor
units
had
actually
decreased
in
size,
as
opposed
to
starting
out
small
and
not
expanding.
Similar
results
were
obtained
for
two
of
the
three
other
muscles
exam-
ined
4-6
weeks
after
partial
denervation.
One
of
these,
partially
dener-
vated
on
day
7
and
examined
on
day
43,
had
two
remaining
motor
units
and
408
large
fibres
visible
in
a
cross-section
of
the
muscle.
The
individual
motor
unit
tensions
were
approximately
equal
in
amplitude
(Text-fig.
8A:
upper
record,
one
motor
unit;
lower
record,
both
motor
units)
but
they
were
several
times
larger
in
absolute
value
than
those
in
a
normal
muscle
of
the
same
age,
owing
in
part
to
the
hypertrophy
of
the
inner-
vated
fibres.
The
total
tension
produced
by
stimulating
the
nerve
close
to
the
muscle
(Text-fig.
8B)
was
no
larger
than
that
from
combined
stimula-
tion
of
the
two
motor
units,
indicating
that
there
were
no
other
large
motor
units
in
the
nerve.
Moreover,
the
tension
from
direct
muscle
stimulation
was
the
same
as
the
nerve-evoked
tension,
suggesting
that
non-innervated
fibres
(presumably
all
the
atrophied
ones)
contributed
little
to
the
total
muscle
tension.
A
low
level
of
multiple
innervation
was
found
in
this
muscle
(four
of
twenty
fibres
tested).
Accordingly,
the
estimated
size
of
each
motor
unit,
based
on
the
muscle
fibre
counts
and
allowing
for
20
%
of
the
large
fibres
being
doubly
innervated,
was
about
245
fibres,
or
8
%
of
the
normal
total.
In
another
moscle,
partially
denervated
on
day
7
and
examined
on
day
41,
only
one
surviving
motor
unit
was
counted
(plus
a
few
very
small
motor
units
that
had
regenerated
through
the
cut
nerve);
we
estimated
that
it
innervated
280
muscle
fibres,
or
9
%
of
the
normal
total.
Finally,
one
muscle
having
six
surviving
motor
axons
was
examined
at
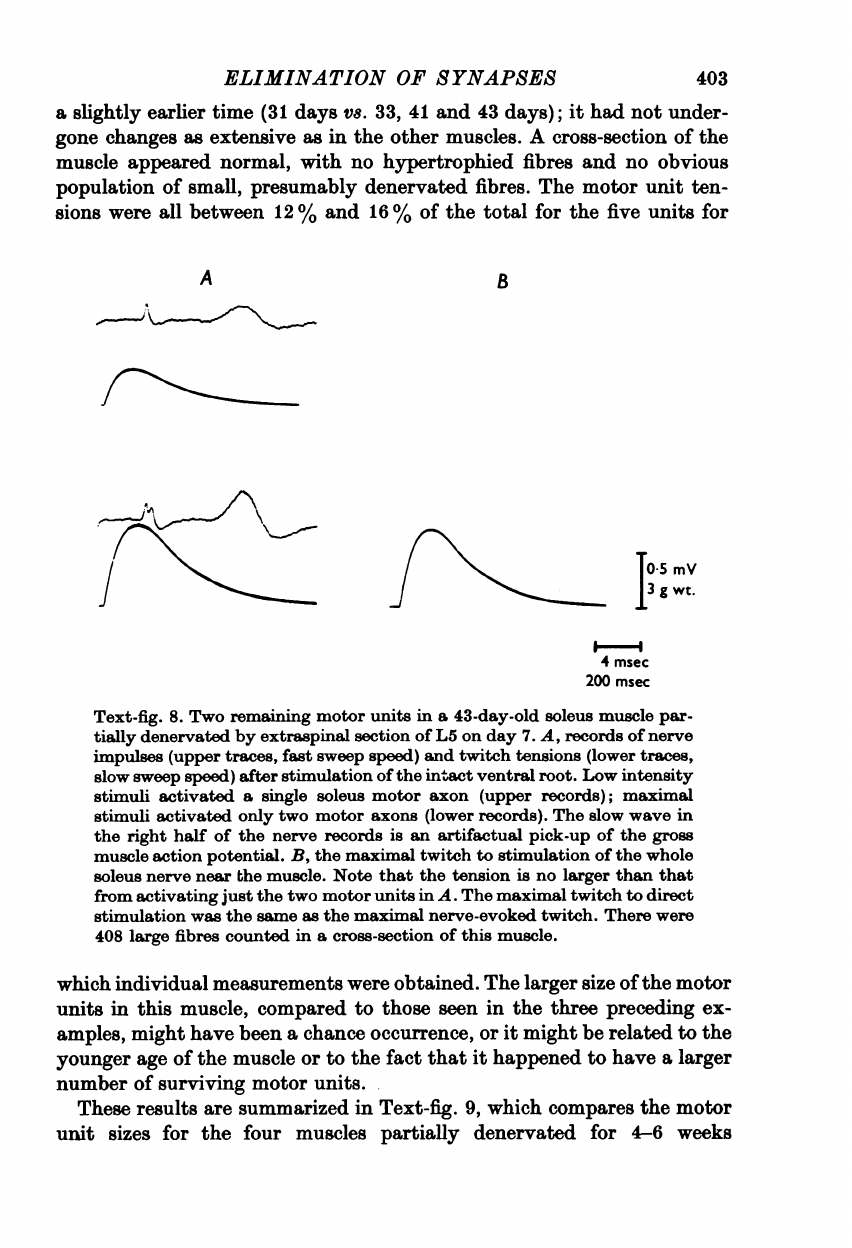
ELIMINATION
OF
SYNAPSES
a
slightly
earlier
time
(31
days
v8.
33,
41
and
43
days);
it
had
not
under-
gone
changes
as
extensive
as
in
the
other
muscles.
A
cross-section
of
the
muscle
appeared
normal,
with
no
hypertrophied
fibres
and
no
obvious
population
of
small,
presumably
denervated
fibres.
The
motor
unit
ten-
sions
were
all
between
12
%
and
16
%
of
the
total
for
the
five
units
for
A
B
An
I
I~~~~~~~~~~~~~~~~~~~3
g
wt.
4
msec
200
msec
Text-fig.
8.
Two
remaining
motor
units
in
a
43-day-old
soleus
muscle
par-
tially
denervated
by
extraspinal
section
of
L5
on
day
7.
A,
records
of
nerve
impulses
(upper
traces,
fast
sweep
speed)
and
twitch
tensions
(lower
traces,
slow
sweep
speed)
after
stimulation
of
the
intact
ventral
root.
Low
intensity
stimuli
activated
a
single
soleus
motor
axon
(upper
records);
maximal
stimuli
activated
only
two
motor
axons
(lower
records).
The
slow
wave
in
the
right
half
of
the
nerve
records
is
an
artifactual
pick-up
of
the
gross
muscle
action
potential.
B,
the
maximal
twitch
to
stimulation
of
the
whole
soleus
nerve
near
the
muscle.
Note
that
the
tension
is
no
larger
than
that
from
activating
just
the
two
motor
units
in
A.
The
maximal
twitch
to
direct
stimulation
was
the
same
as
the
maximal
nerve-evoked
twitch.
There
were
408
large
fibres
counted
in
a
cross-section
of
this
muscle.
which
individual
measurements
were
obtained.
The
larger
size
of
the
motor
units
in
this
muscle,
compared
to
those
seen
in
the
three
preceding
ex-
amples,
might
have
been
a
chance
occurrence,
or
it
might
be
related
to
the
younger
age
of
the
muscle
or
to
the
fact
that
it
happened
to
have
a
larger
number
of
surviving
motor
units.
These
results
are
summarized
in
Text-fig.
9,
which
compares
the
motor
unit
sizes
for
the
four
muscles
partially
denervated
for
4-6
weeks
403

404
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
(continuous
lines)
with
those
seen
around
the
time
of
the
initial
operation
(days
3,
5
and
7,
dashed
lines).
The
mean
size
of
motor
units
was
consid-
erably
lower
in
the
partially
denervated
muscles
(10.6
vs.
15-9
%),
es-
pecially
if
one
considers
only
the
three
muscles
with
an
obvious
mixture
of
hypertrophied
and
atrophied
fibres
(hatched
area,
mean
7
%),
excluding
the
remaining
muscle
(dotted
area)
on
the
grounds
that
its
histological
uniformity
suggests
that
it
might
still
have
been
in
a
transitional
state.
10
0
.......I
I
_I
I
I
I
4J
I
I
0
1
0
3
M
_
~~I
I
e
_
11
.o5
1
z
I
I
0
10
20
30
Motor
unit
sizes
Text-fig.
9.
Histogram
of
unit
sizes
at
late
times
after
partial
denervation.
Hatched
area:
motor
unit
sizes
for
five
units
in
three
soleus
muscles
examined
at
33,
41
and
43
days
after
birth
following
partial
denervation
at
4,
7
and
7
days,
respectively.
The
estimates
were
based
on
counts
of
large,
non-atrophied
muscle
fibres
seen
in
histological
cross-sections,
expressed
as
a
percentage
of
the
normal
value
of
3000
fibres.
Dotted
area:
motor
unit
sizes,
based
on
tension
measurements
for
five
units
in
a
muscle
partially
denervated
on
day
4
and
examined
on
day
31.
Dashes
lines:
histogram
of
motor
unit
sizes
for
twenty-six
motor
units
in
three
normal
muscles
aged
3,
5,
and
7
days
(from
Text-fig.
6).
In
any
event,
whether
or
not
the
one
muscle
is
excluded
from
the
sample,
the
results
are
consistent
with
a
genuine
shrinkage
of
motor
units
during
maturation
after
partial
denervation.
They
are
not
conclusive,
however,
owing
to
the
relatively
large
scatter
in
motor
unit
sizes
and
to
the
uncer-
tainties
that
result
from
comparisons
involving
two
different
methods
for
measuring
motor
unit
sizes
(tension
measurements
and
muscle
fibre

ELIMINATION
OF
SYNAPSES
counts).
We
conclude
that,
at
the
very
least,
there
is
no
widespread
sprouting
of
axon
collaterals
to
take
over
large
numbers
of
denervated
fibres
after
partial
denervation
in
immature
rats.
To
the
contrary,
it
is
prob-
able
that
some
motor
units
are
not
able
to
maintain
their
original
comple-
ment
of
peripheral
synapses
even
though
some
muscle
fibres
consequently
are
stripped
of
their
remaining
inputs.
Re-innervation
and
cro88-innervation
of
immature
mwdcle
By
examining
the
events
that
occur
during
re-innervation
of
the
im-
mature
soleus
muscle
by
its
original
nerve
and
cross-innervation
by
a
foreign
nerve
it
has
been
possible
to
study
several
questions
that
would
be
difficult
or
impossible
to
investigate
in
a
normal
muscle.
For
example,
we
were
interested
in
whether
newly
formed
terminals
can
take
part
in
the
process
of
synapse
elimination,
whether
interactions
between
synapses
situated
at
a
distance
from
one
another
can
lead
to
the
elimination
of
one
of
them
and
whether
there
is
any
selectivity
in
the
choice
of
which
synapses
are
to
survive
and
which
are
to
be
removed.
Re-innervation
by
the
original
nerve
Re-innervation
of
the
immature
soleus
muscle
occurred
relatively
quickly
and
completely
following
a
crush
of
the
nerve
just
at
its
entry
into
the
muscle.
Substantial
contractions
to
nerve
stimulation
were
seen
within
a
week
following
denervation
on
the
first
or
second
day
after
birth.
By
days
11-12
more
than
half
of
the
muscle
had
been
re-innervated,
as
assessed
by
micro-electrode
surveys
of
superficial
muscle
fibres
and
by
comparing
nerve-evoked
and
direct
muscle
tensions
(three
experiments).
Three
weeks
after
birth
re-innervation
was
nearly
complete
(80-100
%
in
two
experiments);
muscle
cross-sections
taken
at
this
time
showed
a
normal
number
of
muscle
fibres
having
relatively
uniform
diameters,
indicating
that
there
had
not
been
a
loss
or
severe
atrophy
of
large
num-
bers
of
fibres.
During
the
first
two
weeks
of
re-innervation
many
muscle
fibres
were
innervated
by
more
than
one
motor
axon.
In
some
muscles
as
many
as
80-90
%
of
the
re-innervated
fibres
had
e.p.p.s
with
more
than
one
com-
ponent.
Text-fig.
10
shows
the
time
course
of
elimination
of
polyneuronal
innervation
in
the
re-innervated
muscles.
Most
of
the
synapses
that
were
eliminated
during
days
10-15
were
presumably
formed
during
the
prece-
ding
week
when
re-innervation
was
taking
place
most
rapidly
and
were
therefore
much
younger
than
synapses
in
normal
muscles,
which
are
formed
in
the
final
week
of
gestation
(Bennett
&
Pettigrew
1974a).
It
is
remarkable
how
closely
the
decline
in
multiple
innervation
followed
that
seen
in
normal
muscles
(dashed
line,
taken
from
Text-fig.
4).
The
405

406
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
elimination
process
seemed
to
run
its
normal
course
even
though
the
synapses
it
acted
upon
were
about
half
their
normal
age.
In
the
great
majority
of
fibres
examined
in
the
re-innervated
muscles
all
components
of
the
e.p.p.
had
similar
rapid
time
courses,
just
as
in
normal
immature
muscles.
This
indicated
that
they
were
all
associated
with
end-
plate
cholinesterase,
and
that
they
were
relatively
close
to
one
another.
Occasional
e.p.p.s.
had
components
of
clearly
different
rise
times,
suggest-
ing
that
they
might
be
spatially
separated.
In
order
to
examine
the
spatial
100
-
0
%
C
75
-
*741
o.S
50
Co
I,
25
_-
o
{~~~
1
._
NO
~~~~~~0
41
U
.00_
O
'
'
J
//L
'^-
' '
5
10
15
20
25
30
50
75
100
Age
(days)
Text-fig.
10.
Incidence
of
multiple
innervation
during
re-innervation
by
the
soleus
nerve.
Each
point
shows
the
percentage
of
re-innervated
fibres
in
which
more
than
one
e.p.p.
component
was
detected.
At
least
twenty
fibres
were sampled
in
each
muscle.
The
soleus
nerve
was
crushed
next
to
its
entry
into
the
muscle
on
the
first
or
second
day
after
birth.
Some
muscles
had
also
been
cross-innervated
through
the
fibular
nerve
but
these
mea-
surements
were
made
in
parts
of
the
muscle
well
away
from
the
zone
of
foreign
innervation.
Note
that
very
occasional
multiple
inputs
were
seen
even
at
long
times
after
birth.
The
interrupted
line
shows
the
time
course
of
elimination
of
multiple
inputs
in
normal
muscles
(from
Text-fig.
4).
distribution
of
synapses
more,
closely
we
looked
at
the
disposition
of
cholinesterase
in
single
muscle
fibres
isolated
from
the
re-innervated
muscles.
All
of
the
end-plates
were
situated
in
the
vicinity
of
the
original
end-plate
band,
presumably
because
the
regenerating
axons
were
guided
there
along
the
old
nerve
branches.
Most
fibres
had
only
a
single
end-plate,
even
in
muscles
where
the
majority
of
fibres
were
multiply
innervated.
For
example,
in
one
10-day-old
re-innervated
muscle,
eighteen
of
twenty
fibres
tested
physiologically
were
multiply
innervated,
whereas
only
eight

ELIMINATION
OF
SYNAPSES
out
of
twenty-one
fibres
isolated
from
the
same
region
of
the
muscle
had
more
than
one
end-plate.
Thus,
it
is
likely
that
some
of
the
regenerating
axons,
converged
on
to
the
same
end-plate
site,
just
as
in
normal
immature
muscle.
This
site
is
probably
the
original
end-plate
present
before
dener-
vation
but
this
is
not
certain
in
view
of
the
lability
of
end-plate
cholin-
esterase
in
immature
fibres
(see
below).
The
presence
of
some
multiple
cholinesterase
spots
(twenty-five
double
sites
andtwo
triple
sites
in
ninety-
two
fibres
from
six
muscles
aged
10
to
21
days)
suggests
that
not
all
axons
returned
exactly
to
the
old
end-plates
and
that
some
fibres
might
have
been
innervated
at
more
than
one
location.
It
was
difficult,
however,
to
estimate
the
frequency
of
occurrence
of
distributed
end-plates
because
of
the
problem
in
knowing
whether
the
old
end-plate
had
been
re-innervated,
whether
it
was
present
but
still
denervated,
or
whether
it
had
been
elimi-
nated.
A
better
understanding
of
the
spatial
distribution
of
newly-formed
synapses
came
from
examining
muscles
that
had
been
cross-innervated
by
a
foreign
nerve
in
a
region
well
outside
the
original
end-plate
zone.
Cross-innervation
by
a
foreign
nerve
Cross-innervation
of
the
immature
soleus
muscle
took
place
only
after
disruption
of
the
original
nerve
supply,
just
as
in
adult
muscle
(Elsberg
1917;
Jansen,
Lomo,
Nicolaysen
&
Westgaard,
1973).
In
three
new-born
rats
the
superficial
branch
of
the
fibular
nerve
was
transplanted
on
to
the
soleus
muscle
without
interrupting
the
original
nerve.
At
the
time
of
the
final
experiment,
nineteen
days
after
birth,
there
were
no
contractions
to
stimulation
of
the
foreign
nerve
in
any
of
the
muscles,
even
though
the
nerve
had
grown
extensively
over
the
surface
of
the
muscle;
nor
were
any
sub-threshold
e.p.p.s
detected
in
the
one
muscle
where
a
micro-electrode
survey
was
made.
In
contrast,
innervation
by
the
foreign
nerve
took
place
readily
when
the
soleus
nerve
was
cut
or
crushed.
As
in
the
adult
(Frank
et
al.
1975)
the
foreign
nerve
never
took
over
more
than
about
a
third
of
the
muscle,
even
when
re-innervation
was
delayed
by
cutting
or
re-secting
the
original
nerve.
Two
types
of
polyneuronal
innervation
involving
the
foreign
nerve
were
seen.
Some
fibres
received
separate
inputs
from
the
foreign
and
original
nerves,
while
other
fibres
received
more
than
one
input
through
the
foreign
nerve
alone.
These
two
forms
of
polyneuronal
innervation
will
be
con-
sidered
separately,
since
our
methods
for
testing
for
them
differed
slightly
and,
more
importantly,
because
there
was
a
striking
difference
in
the
persistence
of the
two
types
of
multiple
innervation
during
maturation.
407

408
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
The
interaction
between
foreign
and
original
nerves
The
extent
of
innervation
by
each
nerve
was
assayed
by
recording
action
potentials
and
e.p.p.s.
intracellularly
from
a
series
of
superficial
muscle
fibres,
noting
whether
each
fibre
was
innervated
by
one,
both
or
neither
nerve.
Twenty-one
muscles
were
examined
at
times
ranging
from
8
to
99
days
after
soleus
nerve
crush
and
fibular
nerve
implant.
The
usual
result
from
this
survey
was
that
the
foreign
nerve
tended
to
innervate
only
the
60
L
*
~~B
C
C
so
.
~~0
L,,
3.0
2
mV
~
~ ~ ~ ~
~~
[
20
20
msec
a
b
0
25
50
75
100
Age
(days)
Text-fig.
11.
Dual
innervation
of
individual
soleus
muscle
fibres
by
both
original
and
transplanted
foreign
(fibular)
nerve.
A,
intracellular
record
of
end-plate
potentials
to
both
soleus
nerve
stimulation
(a)
and
fibular
nerve
stimulation
(b)
from a
9-day-old
animal
in
which
the
superficial
fibular
nerve
was
transplanted
and
the
soleus
nerve
crushed
8
days
before.
B,
per-
centage
of
cross-innervated
fibres
that
were
also
re-innervated
by
the
original
nerve
in
muscles
of
different
ages.
The
initial
operation
was
per-
formed
at
days
1
or
2
in
all
animals.
Each
point
represents
the
observations
in
one
soleus
muscle
in
which
at
least
twenty
cross-innervated
muscle
fibres
were
examined.
lateral
one
third
to
one
half
of
the
muscle,
and
in
this
region
very
few
fibres
had
in
addition
been
re-innervated
by
the
original
nerve.
Fibres
in
the
medial
half
of
the
muscle
were
innervated
only
by
the
soleus
nerve.
In
the
border
region
between
the
two
territories
many
fibres
received
an
input
from
both
nerves.
Text-fig.
11
A
shows
an
example
of
a
fibre
with
a
dual
input
from
the
two
nerves,
taken
from
a
9-day-old
muscle.
Stimula-
tion
of
the
soleus
nerve
evoked
an
e.p.p.
having
two
components
with
slightly
different
latencies
and
stimulation
of
the
fibular
nerve
evoked
an
e.p.p.
of
similar
amplitude.
As
an
index
of
the
frequency
of
dual
innervation
by
the
two
nerves
we
determined
what
percentage
of
muscle
fibres
innervated
by
the
foreign

ELIMINATION
OF
SYNAPSES
nerve
had
in
addition
an
input
from
the
original
nerve.
Text-fig.
11
B
shows
the
results
for
twenty-one
muscles
in
which
ten
to
fifty
fibres
with
foreign
innervation
were
examined
in
each
muscle.
The
percentage
of
dually
innervated
fibres
varied
over
a
wide
range
but
it
did
not
appear
to
change
significantly
with
time.
The
average
percentage
for
muscles
under
25
days
of
age
(19.5
%)
is
not
significantly
different
from
that
for
older
muscles
(18
%),
nor
was
there
a
significant
difference
if
any
other
age
was
chosen
as
the
dividing
point.
The
particular
values
obtained
for
the
per-
centage
of
dually
innervated
fibres
in
each
muscle
may
not
be
very
accurate
owing
to
the
sampling
problems
that
arose
because
these
fibres
tended
to
lie
in
a
narrow
strip
between
the
foreign
and
original
nerve
territories.
Nevertheless,
the
results
demonstrate
that
substantial
numbers
of
fibres
with
both
foreign
and
original
nerve
synapses
can
be
found
at
all
ages,
and
that
there
is
no
dramatic
reduction
in
their
incidence
as
a
function
of
time.
It
is
surprising
that
the
majority
of
fibres
innervated
by
the
foreign
nerve
never
became
re-innervated
through
the
original
nerve.
The
failure
to
re-innervate
cross-innervated
muscle
fibres
in
the
immature
animals
was
not
due
simply
to
a
lowered
over-all
capacity
for
regeneration
by
the
soleus
nerve,
because
in
the
absence
of
the
foreign
nerve
the
soleus
nerve
was
capable
of
completely
re-innervating
its
muscle,
as
mentioned
above.
It
thus
seems
that
the
presence
of
foreign
innervation
can
inhibit
re-
innervation
of
old
end-plates
in
immature
rats.
The
separation
between
foreign
and
original
synapses
on
dually-
innervated
fibres
was
determined
by
staining
the
muscle
for
end-plate
cholinesterase.
In
all
but
one
experiment
the
region
of
foreign
end-plates
was
in
the
proximal
part
of
the
muscle,
well
away
from
the
original
end-
plate
band,
just
as
is
seen
in
adult
soleus
muscles
after
cross-innervation
(Frank
et
al.
1976).
The
minimal
separation
between
the
original
end-plate
band
and
the
closest
foreign
end-plates
was
at
least
1
mm
and
usually
more.
In
the
one
exceptional
case
the
foreign
nerve
had
grown
directly
into
the
original
end-plate
region
and
made
synapses
there.
It
is
perhaps
significant
that
this
was
the
one
muscle
in
which
we
did
not
find
any
fibres
receiving
an
input
from
both
foreign
and
original
nerves.
There
was
usually
a
sharp
transition
in
the
appearance
of
the
original
end-plate
band
just
at
the
border
between
foreign
and
original
nerve
territories.
The
band
was
continuous
and
of
normal
appearance
in
the
re-
innervated
zone
while
it
was
much
less
dense
and
consisted
of
scattered
isolated
end-plates
in
that
part
of
the
muscle
which
was
cross-innervated
by
the
foreign
nerve.
Examination
of
single,
isolated
fibres
confirmed
that
the
original
end-plate
cholinesterase
had
often
completely
disappeared
from
cross-innervated
fibres.
On
those
fibres
which
had
both
foreign
and
409

410
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
original
end-plates
the
distance
between
them
was,
as
expected,
always
1
mm
or
more.
The
disappearance
of
the
cholinesterase
at
the
original
end-plates
was
not
simply
a
consequence
of
the
end-plate
itself
not
being
re-innervated.
Following
simple
denervation
of
neonatal
muscles
cholinesterase
remains
at
the
original
end-plate
for
at
least
a
month
(Lubinska
&
Zelena
1966).
In
contrast,
we
found
that
the
cholinesterase
at
denervated
end-plates
on
fibres
innervated
elsewhere
by
a
foreign
nerve
disappeared
entirely
within
about
a
week.
A
clear
illustration
of
this
came
from
a
13-day-old
muscle
in
which
the
fibular
nerve
had
been
implanted
on
the
first
day
after
birth
and
the
soleus
nerve
cut
far
enough
from
the
muscle
to
prevent
re-
innervation
by
the
time
of
the
final
experiment.
In
the
region
of
the
muscle
innervated
by
the
foreign
nerve
only
scattered
end-plates
were
visible
in
the
original
end-plate
band,
whereas
outside
the
region
of
cross-innerva-
tion
the
old
denervated
end-plates
appeared
in
the
whole
mount
as
a
continuous
dense
band
of
cholinesterase.
Isolation
of
individual
fibres
teased
from
the
cross-innervated
region
of
the
muscle
demonstrated
that
seven
out
of
eight
fibres
having
foreign
nerve
end-plates
had
completely
lost
their
original
end-plates;
the
one
fibre
having
both
foreign
and
original
end-plates
presumably
had
been
cros-innervated
for
a
shorter
time
than
the
others.
We
did
not
investigate
in
detail
the
time
required
for
foreign
innervation
to
suppress
cholinesterase
at
the
original
end-plate
but
it
is
clear
that
the
process
cannot
take
much
more
than
a
week.
The
lability
of
end-plate
cholinesterase
indicated
by
this
experiment
might
be
related
to
the
inability
of
regenerating
soleus
nerve
axons
to
re-innervate
most
cross-
innervated
muscle
fibres.
Polyneuronal
innervation
through
the
foreign
nerve
At
early
times
after
cross-innervation,
graded
stimulation
of
the
fibular
nerve
showed
that
many
muscle
fibres
were
innervated
by
more
than
one
foreign
motor
axon.
Two
lines
of
evidence
indicate
that
many
of
the
foreign
synapses
on
multiply
innervated
fibres
were
situated
at
an
appreciable
distance
from
one
another.
First,
the
different
components
of
the
e.p.p.
often
had
clearly
different
time
courses.
For
example,
in
the
fibre
illus-
trated
in
Text-fig.
12A
a
low
intensity
stimulus
(left)
to
the
fibular
nerve
elicited
an
e.p.p.
with
a
rise
time
of
about
12
msec,
while
a
stronger
stim-
ulus
(right)
evoked
an
additional
component
whose
rise
time
was
5
msec.
A
likely
explanation
for
the
difference
in
time
course
is
that
the
two
synapses
were
located
at
different
distances
from
the
recording
micro-electrode.
More
direct
evidence
for
spatially
separated
end-plates
came
from
exam-
ining
the
distribution
of
cholinesterase
on
individual
fibres
isolated
from

ELIMINATION
OF
SYNAPSES
411
the
region
of
foreign
innervation.
Many
fibres
had
two
distinct
end-plates
separated
by
distances
of
up
to
1
mm;
an
example
of
a
fibre
having
two
end-plates
about
150
,um
apart
is
shown
in
P1.
20
(arrows).
A
few
fibres
had
three
separate
end-plates
in
the
region
of
the
fibular
nerve,
and
on
one
fibre
five
closely
spaced
but
distinct
end-plates
were
found.
Multiple
fibular
end-plates
were
seen
in
more
than
a
quarter
of
the
fibres
isolated
from
four
cross-innervated
muscles
between
10
and
13
days
old
(thirty
out
of
107
fibres).
In
the
same
four
muscles
the
incidence
of
polyneuronal
innervation
estimated
physiologically
was
about
one
fibre
in
three
(twenty-
eight
of
seventy-nine
fibres).
This
suggests
that
most
but
perhaps
not
all
polyneuronal
innervation
mediated
by
the
foreign
nerve
occurs
at
spa-
tially
separate
synapses.
A
100
_
B
0>
75
"C
4)0
LLJ
50
msec
0
5
10
15
20
25
3050
75100
Age
(days)
Text-fig.
12.
Polyneuronal
innervation
by
a
foreign
nerve.
A,
multiple
e.p.p.
components
evoked
by
fibular
nerve
stimulation
in
a
19-day-old
soleus
muscle
cross-innervated
on
day
2.
Low
intensity
stimuli
(left)
produced
a
small
e.p.p.
with
rise
time
about
twice
as
long
as
that
of
the
higher
threshold
component
(right).
B,
percentage
of
foreign
innervated
fibres
in
which
more
than
one
e.p.p.
component
was
detected
during
graded
stimulation
of
the
foreign
nerve.
Each
point
gives
the
results
for
one
muscle
in
which
at
least
ten
and
usually
twenty
or
more
cross-innervated
fibres
were
ex-
amined.
The
interrupted
line
shows
the
time course
of
synapse
elimination
in
normal
muscles
(from
Text-fig.
4).
All
physiological
signs
of
polyneuronal
innervation
through
the
foreign
nerve
eventually
disappeared
in
the
cross-innervated
muscles.
Text-fig.
12B
shows
the
time
course
of
elimination
of
multiple
innervation
as
deter-
mined
physiologically
with
intracellular
recordings.
Each
point
represents
the
observations
in
one
muscle
and
gives
the
percentage
of
fibres
with
more
than
one
input
from
the
fibular
nerve.
The
incidence
of
polyneuronal
in-
nervation
was
quite
high
up
to
about
the
tenth
day
after
birth,
with
80
%
or
more
multiple
innervation
in
some
muscles.
The
disappearance
of
multiple
innervation
initially
was
similar
to
that
seen
in
normal
muscles
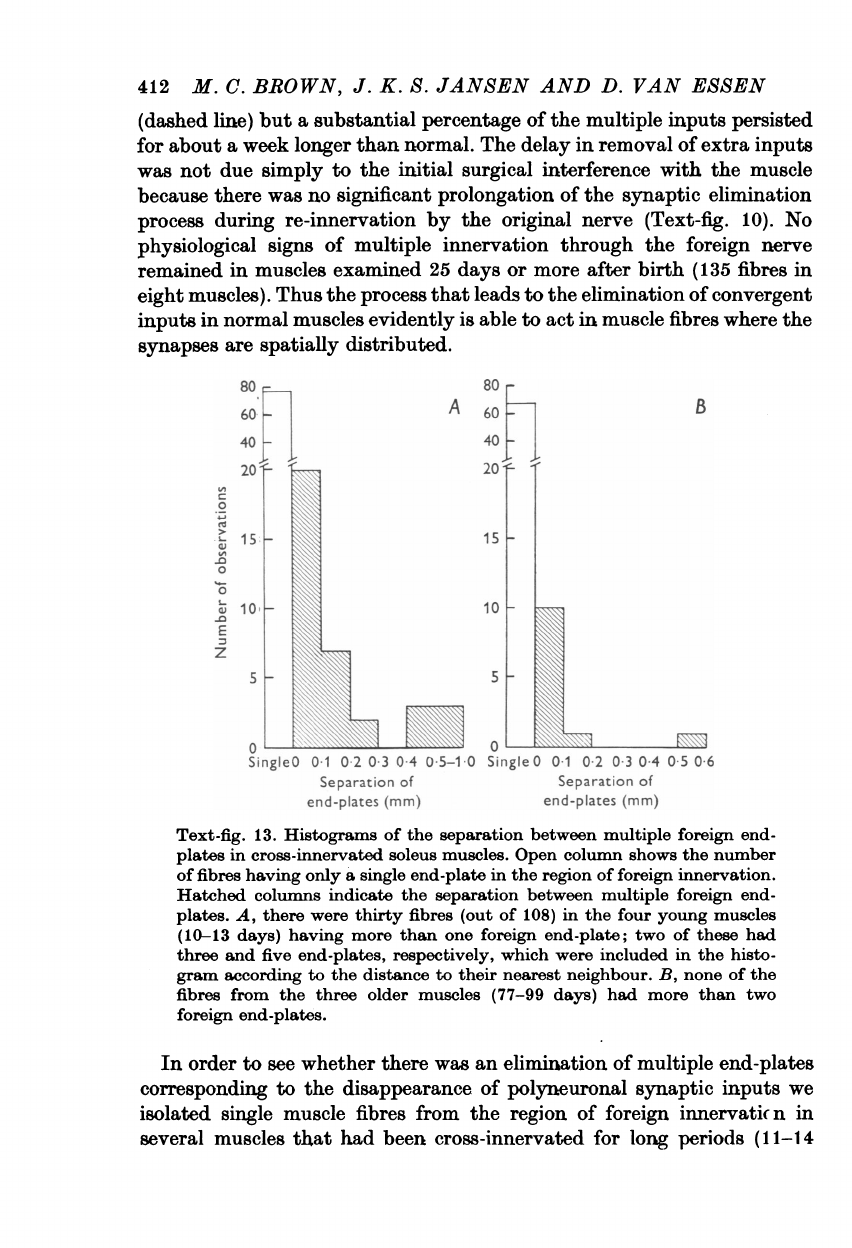
412
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
(dashed
line)
but
a
substantial
percentage
of
the
multiple
inputs
persisted
for
about
a
week
longer
than
normal.
The
delay
in
removal
of
extra
inputs
was
not
due
simply
to
the
initial
surgical
interference
with
the
muscle
because
there
was
no
significant
prolongation
of
the
synaptic
elimination
process
during
re-innervation
by
the
original
nerve
(Text-fig.
10).
No
physiological
signs
of
multiple
innervation
through
the
foreign
nerve
remained
in
muscles
examined
25
days
or
more
after
birth
(135
fibres
in
eight
muscles).
Thus
the
process
that
leads
to
the
elimination
of
convergent
inputs
in
normal
muscles
evidently
is
able
to
act
in
muscle
fibres
where
the
synapses
are
spatially
distributed.
so
s
20
2
A
60
B
60
40
40
-
is
.0
0
0
10.
-
~~~~~10
E
z
55
0
~~~~~~~0
SingleO
0-1
0-2
0-3
04
05-10
Single
0
0-1
02
03
0-4
0-5
0-6
Separation
of
Separation
of
end-plates
(mm)
end-plates
(mm)
Text-fig.
13.
Histograms
of
the
separation
between
multiple
foreign
end-
plates
in
cross-innervated
soleus
muscles.
Open
column
shows
the
number
of
fibres
having
only
a
single
end-plate
in
the
region
of
foreign
innervation.
Hatched
columns
indicate
the
separation
between
multiple
foreign
end-
plates.
A,
there
were
thirty
fibres
(out
of
108)
in
the
four
young
muscles
(10-13
days)
having
more
than
one
foreign
end-plate;
two
of these
had
three
and
five
end-plates,
respectively,
which
were
included
in
the
histo-
gram
according
to
the
distance
to
their
nearest
neighbour.
B,
none
of
the
fibres
from
the
three
older
muscles
(77-99
days)
had
more
than
two
foreign
end-plates.
In
order
to
see
whether
there
was
an
elimination
of
multiple
end-plates
corresponding
to
the
disappearance
of
polyneuronal
synaptic
inputs
we
isolated
single
muscle
fibres
from
the
region
of
foreign
innervation
in
several
muscles
that
had
been
cross-innervated
for
long
periods
(11-14

ELIMINATION
OF
SYNAPSES
weeks)
and
compared
the
results
with
those
obtained
from
younger
muscles
(10-13
days).
It
was
surprising
to
find
that
despite
a
clear
reduc-
tion
in
the
incidence
of
spatially
separated
end-plates
compared
with
that
seen
in
younger
muscles,
there
was
still
a
substantial
number
of
fibres
that
retained
more
than
one
foreign
end-plate.
Text-fig.
13
shows
histograms
of
the
distance
between
multiple
foreign
end-plates
in
young
muscles
(A)
and
in
older
muscles
(B).
Almost
all
of
the
double
end-plates
that
persisted
in
the
older
muscles
were
within
100
,um
of
one
another
(e.g.
P1.
2D),
whereas
many
of
the
end-plates
in
the
younger
muscles
were
separated
by
distances
up
to
1
mm.
The
reduction
in
the
incidence
of
end-
plates
separated
by
more
than
100
#tm
provides
evidence
that
the
process
of
synapse
elimination
can
work
on
synapses
that
were
sufficiently
mature
to
have
induced
the
incorporation
of
cholinesterase
at
a
new
end-plate.
DISCUSSION
The
location
of
synapses
on
immature
muscle
fibres
The
argument
that
each
immature
muscle
fibre
receives
a
direct
and
convergent
input
from
more
than
one
motor
neurone
depends
upon
several
lines
of
evidence,
none
of
which
is
by
itself
conclusive.
Our
electro-
physiological
experiments
show
that
all
of
the
synaptic
inputs
to
any
particular
fibre
are
at
most
about
50
/tm
apart
(from
the
desensitizing
effects
of
iontophoretically
applied
ACh),
and
that
all
of
the
synapses
must
be
associated
with
end-plate
cholinesterase
(from
the
effects
of
prostig-
mine
on
e.p.p.
time
course).
Since
there
is
only
one
cholinesterase
spot
per
immature
muscle
fibre,
it
follows
that
either
the
multiple
inputs
all
con-
verge
on
to
a
single
end-plate,
or
that
there
is
strong
electrical
coupling
between
neighbouring
fibres
whose
end-plates
are
immediately
adjacent
to
one
another.
We
found
no
sign
of
widespread
electrical
coupling
between
immature
muscle
fibres,
although
the
possibility
was
not
excluded
that
there
might
be
coupling
within
small
groups
of
fibres
linked
closely
enough
to
appear
as
a
single
fibre
in
the
dissecting
microscope.
On
the
other
hand,
each
end-plate
in
an
immature
muscle
is
supplied
by
several
preterminal
axons
and
all
but
one
of
these
disappear
during
maturation.
Although
we
could
not
show
directly
that
each
preterminal
branch
belonged
to
a
differ-
ent
motor
axon,
it
seems
very
likely
that
the
multiple
preterminal
axons
are
the
anatomical
basis
for
the
presence
of
several
e.p.p.
components
in
each
immature
muscle
fibre.
Finally,
it
is
clear
that
the
multiple
synaptic
inputs
demonstrated
by
graded
stimulation
of
the
nerve
do
not
come
about
because
of
a
division
of
each
motor
axon
into
several
separate
branches
that
subsequently
converge
on
to
the
same
set
of
muscle
fibres.
This
is
because
the
tensions
produced
by
stimulating
individual
motor
units
in
413
PH
Y
26i
I4

414
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
ventral
root
filaments
are
so
large
that
there
must
inevitably
be
a
large
overlap
in
the
innervation
of
different
motor
units.
We
therefore
conclude,
in
agreement
with
Redfern
(1970)
and
Bennett
&
Pettigrew
(1974a)
that
there
is
genuine
polyneuronal
innervation
in
immature
skeletal
muscles
of
the
rat.
In
muscles
that
are
only
a
few
days
old
all
muscle
fibres
receive
at
least
two
inputs
and
estimates
of
the
average
degree
of
multiple
innerva-
tion
range
from
about
three
(Bennett
&
Pettigrew,
1974a)
to
about
five
(Text-fig.
6,
above).
It
is
remarkable
that
all
of
the
inputs
are
crowded
into
an
area
that
is
smaller
than
an
end-plate
on
an
adult
muscle
fibre,
yet
each
synapse
is
able
to
excite
the
muscle
fibre
on
its
own.
Presumably
the
efficiency
of
transmission
is
related
in
part
to
the
higher
input
resistance
of
smaller
muscle
fibres.
Polyneuronal
innervation
has
also
been
seen
in
the
soleus
muscle
of
kittens
(Bagust
et
al.
1973),
suggesting
that
it
may
be
a
widespread
or
even
universal
event
in
the
maturation
of
mammalian
skeletal
muscle.
The
extent
of
hyperinnervation
which
exists
in
new-born
animals
will,
how-
ever,
presumably
vary
according
to
the
degree
of
maturity
at
birth.
It
will
be
interesting
to
know
whether
in
a
particular
species
the
loss
of
poly-
neuronal
innervation
occurs
synchronously
in
all
muscles, as
is
the
case
for
the
soleus
and
diaphragm
muscles
of
the
rat.
The
elimination
of
synapses
during
normal
maturation
During
the
period
when
polyneuronal
innervation
disappears
the
num-
ber
of
motor
axons
to
the
soleus
muscle
remains
constant,
while
the
aver-
age
size
of
motor
units
decreases
severalfold.
The
elimination
of
multiple
innervation
and
the
reduction
in
motor
unit
size
conceivably
might
come
about
by
muscle
fibres
dividing
in
such
a
way
as
to
leave
only
one
synapse
on
each
fibre.
Chiakulas
&
Pauly
(1965)
have
shown
that
there
may
be
an
increase
of
about
50
%
in
the
number
of
soleus
muscle
fibres
after
the
first
post-natal
week.
They
suggest
that
the
new
fibres
originate
from
myoblasts
rather
than
from
division
of
pre-existing
fibres
but
even
if
some
division
took
place
it
would
be
quantitatively
insufficient
to
account
fully
for
the
loss
of
multiple
innervation.
Hence,
it
is
evident
that
many
terminals
must
eventually
disappear
by
undergoing
either
degeneration
or
retrac-
tion.
H.
Korneliussen
(personal
communication)
found
no
obvious
signs
of
terminal
degeneration
in
electron
micrographs
of
synapses
from
immature
muscles,
suggesting
that
simple
retraction
of
extra
synapses
is
the
more
likely
alternative.
It
would
be
interesting
to
know
whether
e.p.p.
components
that
are
particularly
small
relative
to
other
components
on
the
same
fibre,
or
which
have
a
particularly
long
latency
following
nerve
stimulation
(e.g.
Fig.
2
in
Redfern,
1970)
might
be
in
the
process
of
being
eliminated
but
it is
difficult
to
see
how
to
test
these
possibilities

ELIMINATION
OF
SYNAPSES
directly.
There
are
numerous
other
changes
in
muscle
and
motor
nerve
properties
that
take
place
during
the
time
that
neuromuscular
connexions
are
maturing.
In
addition
to
growing
in
length
and
cross-sectional
area
the
muscle
develops
fast,
or
slow
contractile
properties,
or
both
(Close,
1964),
and
the
nerve
becomes
myelinated
and
gains
in
conduction
velocity.
It
is
not
yet
known
whether
the
removal
of
multiple
innervation
is
specifically
dependent
on
any
of
these
changes.
The
total
spread
of
motor
unit
sizes
within
a
given
muscle
was
con-
siderably
greater
in
young
animals
than
in
the
adult,
with
a
few
motor
units
being
within
the
adult
range
at
all
ages
we
examined.
This
suggests
that
the
shrinkage
of
motor
units
does
not
come
about
simply
by
the
synchronous
loss
by
each
motor
neurone
of
a
fixed
percentage
of
its
peripheral
terminals.
Either
there
must
be
considerable
variability
in
the
times
at
which
different
motor
units
reach
their
peak
size,
or
else
some
motor
units
never
become
much
larger
than
their
eventual
adult
size.
In
either
case,
it
appears
that
at
any
given
time
the
elimination
process
acts
predominantly
on
the
terminals
of
the
largest
motor
units.
This
could
come
about
if
the
terminals
of
small
motor
units
somehow
had
a
selective
advantage
over
the
terminals
of
larger
ones.
One
can
imagine,
for
example,
that
as
a
motor
neurone
gradually
lost
some
of
its
synapses
it
might
be
able
to
supply
its
remaining
terminals
with
some
material
or
quality
that
improved
their
chances
for
survival.
Synapse
elimination
following
partial
denervation
The
removal
of
all
but
a
few
motor
axons
to
the
soleus
muscle
leaves
the
remaining
neurones
with
the
opportunity
to
innervate
large
numbers
of
muscle
fibres
while
facing
only
minimal
competition
at
the
level
of
peri-
pheral
synaptic
interactions.
One
might
expect
to
find
that
whatever
residual
polyneuronal
innervation
was
present
would
be
removed
nor-
mally,
while
the
remaining
motor
units
would
either
remain
constant
in
size
or
even
expand
by
sprouting
on
to
neighbouring
denervated
fibres.
It
thus
came
as
a
surprise
to
find
that
partial
denervation
resulted,
on
the
one
hand,
in
a
delay
in
the
removal
of
polyneuronal
innervation
among
sur-
viving
motor
units
and
that,
on
the
other
hand,
a
reduction
in
motor
unit
size
nevertheless
eventually
took
place.
It
is
puzzling
that
multiple
innervation
should
persist
on
some
muscle
fibres
just
because
most
of
the
motor
input
to
the
muscle
has
been
re-
moved.
The
delay
might
be
an
indirect
effect
of
the
surgical
interference
within
the
spinal
cord
but
this
seems
unlikely
because
multiple
innervation
disappeared
with
its
normal
time
course
in
the
control
soleus
muscle
on
the
opposite
side.
A
more
interesting
possibility
is
that
the
delay
might
be
related
to
the
large
size
of
the
remaining
motor
units:
for
example,
if
the
14-2
415

416
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
maturation
of
synaptic
terminals
proceeded
more
slowly
in
larger
motor
units
and
if
the
process
of
synapse
elimination
on
each
fibre
did
not
begin
until
the
favoured
terminal
reached
a
certain
absolute,
rather
than
relative
maturity,
then
one
would
expect
partial
denervation
to
delay
the
removal
of
multiple
innervation.
The
eventual
reduction
in
motor
unit
size
that
took
place
after
partial
denervation
was
usually
associated
with
gross
hypertrophy
of
the
remain-
ing
innervated
fibres
and
atrophy,
or
at
least
failure
to
grow,
of
the
dener-
vated
fibres.
Many
motor
units
were
closer
to
their
normal
adult
level
than
to
their
mean
initial
size;
this
happened
too
frequently
to
be
accounted
for
by
a
chance
selection
of
small
motor
units
at
the
time
of
the
initial
operation.
Moreover,
it
is
difficult
to
account
for
the
shrinkage
simply
in
terms
of
the
elimination
of
residual
multiple
innervation,
since
small
motor
units
were
seen
in
muscles
having
only
one
or
two
motor
units
left.
Hence
it
seems
likely
that
the
withdrawal
of
synapses
can
continue
even
though
many
previously
innervated
fibres
actually
become
denervated.
It
is
not
surprising
that
there
should
be
a
natural
limit
to
how
many
muscle
fibres
a
motor
neurone
can
innervate.
What
is
interesting,
though,
is
that
there
should
be
an
actual
reduction
in
the
upper
limit
of
motor
unit
size
during
maturation.
This
might
come
about
if
the
total
synaptic
area
that
a
motor
neurone
could
supply
grew
more
slowly
than
the
mean
area
of
individual
end-plates.
Presumably
the
natural
tendency
of
each
motor
unit
to
reduce
its
initial
size
plays
a
major
role
in
the
elimination
of
hyper-
innervation.
It
cannot
be
the
sole
mechanism
involved,
however,
because
there
must
be
some
way
of
insuring
that
each
muscle
fibre
ends
up
with
one
and
only
one
synapse.
There
is
no
inconsistency
between
the
occurrence
of
axonal
sprouting
after
partial
denervation
of
adult
muscles
(Edds,
1953;
Guth,
1962)
and
our
failure
to
detect
sprouting
of
immature
motor
units.
The
lack
of
sprouting
in
immature
muscles
is
presumably
related
to
the
large
initial
size
of
motor
units.
The
existence
of
an
upper
limit
to
motor
neurone
size
is
entirely
compatible
with
a
moderate
degree
of
sprouting
in
the
adult.
Our
results
suggest
that
the
limiting
size
of
mature
motor
units
might
be
substantially
larger
(i.e.
a
factor
of
two
or
so)
than
their
normal
adult
size.
The
relative
uniformity
in
adult
motor
unit
sizes
seen
in
the
soleus
muscle
is
not
found
in
other
muscles
such
as
the
fast-twitch
extensor
digitorum
longus,
where
there
is
about
a
tenfold
range
in
size
(Close,
1967).
It
would
be
interesting
to
know
whether
this
greater
variability
is
related
to
larger
inherent
differences
among
fast
motor
neurones.

ELIMINATION
OF
SYNAPSES
The
elimination
of
synapses
after
re-innervation
and
cross-innervation
Multiple
innervation
through
regenerated
axons
of
the
original
nerve
disappeared
with
a
time
course
similar
to
that
seen
in
normal
muscles.
This
shows
that
the
process
of
synapse
elimination
is
not
heavily
influenced
by
either
the
age
of
the
terminals
involved
or
the
period
of
muscle
inactivity
before
re-innervation
took
place.
The
lack
of
an
effect
is
remarkable
in
view
of
the
delay
in
the
removal
of
hyperinnervation
following
several
other
procedures
that
might
seem
to
be
less
disruptive
to
the
synapses
involved.
For
example,
partial
denervation
leads
to
a
prolonged
innerva-
tion
overlap
among
motor
units
that
were
not
directly
affected
by
the
operation.
In
addition
Benoit
&
Changeux
(1975)
have
shown
that
teno-
tomy
causes
a
delay
in
the
removal
of
polyneuronal
innervation,
and
we
have
seen
a
similar
effect
following
the
injection
of
botulinum
toxin
into
immature
muscles
(M.
C.
Brown,
J.
K.
S.
Jansen
and
D.
Van
Essen,
un-
published
observations).
The
events
that
took
place
during
cross-innervation
by
a
foreign
nerve
differed
in
several
important
respects
from
those
that
occurred
during
re-
innervation
and
during
the
maturation
of
normal
muscles.
Firstly,
many
synapses
on
fibres
multiply
innervated
through
the
foreign
nerve
were
situated
at
separate
sites
up
to
k/mm
apart.
Distributed
synapses
were
never
seen
in
normal
muscles,
anid
were
probably
not
the
most
common
form
of
multiple
innervation
during
re-innervation.
Presumably,
the
difference
between
re-innervation
and
cross-innervation
is
that
regenera-
ting
axons
from
the
original
nerve
tend
to
be
guided
back
to
the
sur-
viving
old
end-plates.
Secondly,
a
moderate
level
of
polyneuronal
inner-
vation
through
the
foreign
nerve
persisted
through
the
third
week
after
birth.
That
these
multiple
inputs
were
event
ally
eliminated
means
that
the
interactions
leading
to
synapse
elimination
can
act
over
considerable
distances.
The
delay
in
synapse
removal
suggests
that
the
process
might
take
longer
when
the
synapses
are
some
distance
apart.
Indirect
support
for
this
comes
from
the
observation
that
there
was
a
higher
incidence
of
fibres
having
e.p.p.
components
with
different
rise
times
in
the
2-3-week-
old
cross-innervated
muscles
than
in
the
younger
ones.
s
final
major
difference
seen
after
foreign
innervation
was
that
some
multiple
end-plates
persisted
indefinitely
in
cross-innervated
muscles.
This
came
about
in
two
distinct
ways:
some
fibres
received
separate
functional
inputs
from
the
foreign
and
original
nerves;
while
other
fibres
had
two
closely
spaced
end-plates
in
the
region
of
foreign
innervation,
even
though
they
had
only
one
input
that
could
be
demonstrated
physiologically.
The
distance
between
synapses
from
the
foreign
and
original
nerves
was
al-
ways
1
mm
or
more,
suggesting
that
the
degree
of
separation
between
417

418
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
synapses
may
determine
whether
one
of
them
will
be
eliminated.
If
dis-
tance
is
indeed
an
important
factor,
then
the
basis
for
synapse
elimination
cannot
be
a
process
that
affects
the
entire
muscle
fibre
equally,
such
as
the
action
potential
or
contractile
activity.
It
is
interesting
that
the
distance
of
about
1
mm
suggested
by
the
present
experiments
is
similar
to
the
spacing
between
synapses
in
slow
muscles
of
the
adult
chicken.
The
development
of
the
adult
pattern
of
innervation
in
the
chicken
appears
to
come
about
by
a
selective
elimination
of
syn-
apses
which
initially
are
more
closely
spaced
than
in
the
adult
(Bennett
&
Pettigrew,
1974b).
Slow
muscles
in
the
chicken
do
not
conduct
action
potentials
and
it
has
been
suggested
that
the
decremental
spread
of
syn-
aptic
potentials
might
be
involved
in
determining
the
spacing
between
synapses
(Gordon,
Perry,
Tuffery
&
Vrbova,
1974).
This
cannot,
however,
be
the
sole
explanation
for
mammalian
skeletal
muscles,
which
conduct
action
potentials
throughout
the
period
when
synapse
elimination
takes
place.
The
persistence
of
double
esterase-stained
end-plates
in
the
region
of
foreign
innervation
appears
to
be
quite
a
different
phenomenon
than
the
dual
innervation
through
foreign
and
original
nerves
that
was
seen
in
the
same
muscles.
Foreign
and
original
end-plates
on
the
same
muscle
fibre
remained
functional
indefinitely,whereas
we
never
saw
physiological
signs
of
inputs
from
more
than
one
foreign
motor
axon
at
long
times
after
cross-
innervation.
If
there
were
functional
synapses
at
both
foreign
end-plates,
they
must
both
have
belonged
to
the
same
parent
axon.
This
is
an
intriguing
possibility,
because
it
suggests
that
a
muscle
fibre
is
not
inherently
resistant
to
maintaining
more
than
one
closely-spaced
end-
plate,
provided
that
they
are
all
supplied
by
one
axon.
The
key
to
protection
of
multiple
synapses
from
elimination
might
be
the
synchrony
in
the
patterns
of
synaptic
activity,
or
it
might
be
some
other
specific
factor
common
to
all
terminals
of
a
particular
motor
neurone.
Differences
between
immature
and
adult
muscle
There
are
several
important
differences
in
the
events
that
take
place
during
re-innervation
and
cross-innervation
of
immature
v8.
adult
muscles.
Perhaps
the
most
striking
are
the
changes
at
the
original
end-plate
region.
Our
results
show
that
immature,
cross-inervated
muscle
fibres,
but
not
fibres
which
are
simply
denervated,
lose
the
cholinesterase
at
the
original
end-plate
quickly
and
completely.
Associated
with
this
is
a
loss
in
most
cross-innervated
fibres
of
the
ability
to
accept
re-innervation
by
the
original
nerve.
In
adult
cross-innervated
soleus
muscle
fibres
the
original
end-plate
properties
are
much
more
stable.
The
intensity
of
cholinesterase
staining
declines
but
never
disappears
and
original
end-plates
will
accept

ELIMINATION
OF
SYNAPSES
re-innervation
for
many
weeks
in
spite
of
foreign
innervation
of
the
same
fibres
(Frank
et
al.
1975).
The
degree
of
polyneuronal
innervation
seen
at
various
stages
of
re-
and
cross-innervation
is
also
different
for
immature
and
adult
rats.
Multiple
innervation
was
seen
in
only
a
small
percentage
(10-15
%)
of
fibres
examined
in
the
first
week
of
re-innervation
of
the
adult
rat
dia-
phragm
(Jansen
&
Van
Essen,
1975).
It
is
not
known
whether
the
multiple
inputs
were
spatially
distributed
and
whether
they
would
eventually
be
removed.
In
adult
cross-innervated
muscles,
polyneuronal
innervation
through
the
foreign
nerve
occurs
commonly,
even
many
months
after
the
operation
(Frank
et
al.
1975).
However,
recent
experiments
by
T.
Lomo
and
C.
R.
Slater
and
by
H.
Sommerschild
(personal
communications)
suggest
that
there
is
a
removal
of
closely
spaced
foreign
synapses
even
in
the
adult
rat.
Possible
mechanism
of
synapse
elimination
The
elimination
of
peripheral
synapses
could
in
principle
be
controlled
by
a
variety
of
different
mechanisms
involving
any
combination
of
the
three
cell
types
present
at
the
neuromuscular
junction:
neurones,
Schwann
cells
and
muscle
fibres.
Our
results
serve
to
restrict
the
number
of
accep-
table
hypotheses
by
specifying
more
explicitly
the
conditions
under
which
synapse
elimination
can
or
cannot
take
place.
The
most
important
res-
traints
are
the
following:
(1)
the
end
result
of
maturation
is
invariably
that
each
muscle
fibre
is
innervated
by
a
single
motor
axon;
(2)
during
normal
maturation
there
are
never
a
substantial
number
of
muscle
fibres
that
are
completely
denervated
but
after
partial
denervation
withdrawal
of
terminals
continues
even
though
some
muscle
fibres
consequently
are
left
without
innervation;
(3)
terminals
from
small
motor
units
are
more
likely
to
survive
than
those
of
larger
units;
(4)
the
elimination
process
has
a
restricted
range:
it
can
act
over
distances
of
hundreds
of
microns
but
not
over
millimetres.
The
elimination
of
synapses
does
not
appear
to
be
a
random
process,
either
in
the
sense
that
a
particular
motor
neurone
loses
a
randomly
selected
proportion
of
its
terminals
(since
this
would
lead
to
transient
denervation
of
many
fibres),
or
in
the
sense
that
on
each
muscle
fibre
there
is
a
random
choice
of
which
terminal
is
to
survive
(since
terminals
of
smaller
units
survive
preferentially).
Instead,
there
seems
to
be
some
sort
of
competition
among
the
synapses
on
each
fibre.
It
may
be
of
interest
to
speculate
on
the
cellular
mechanisms
that
might
operate
in
a
way
con-
sistent
with
these
restraints.
Selection
by
Schwann
cells.
Since
all
of
the
terminals
on
a
normal
im-
mature
muscle
fibre
are
crowded
into
a
single
end-plate,
it
is
possible
to
419

420
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
imagine
that
a
single
Schwann
cell,
or
row
of
Schwann
cells,
could
select
one
of
the
terminals
to
be
the
survivor.
This
could
be
accomplished
by
the
myelination
of
only
one
preterminal
axon,
providing
that
the
remaining
unmyelinated
axons
were
eventually
withdrawn.
The
selection
of
which
axon
to
myelinate
could
be
made
on
the
basis
of
some
quality
such
as
axon
diameter,
in
which
case
the
smaller
motor
units
might
attain
a
selective
advantage
simply
because
their
terminals
become
larger.
The
myelination
hypothesis
cannot,
however,
account
for
the
events
related
to
the
elimination
of
spatially
separated
end-plates
in
any
simple
way.
Interactions
between
nerve
terminate.
The
elimination
process
might
in-
volve
interactions
only
between
the
nerve
terminals
co-existing
on
a
fibre,
without
any
direct
participation
of
the
muscle
fibre
or
of
Schwann
cells.
The
interactions
might
take
the
form
of
one
terminal
physically
crowding
out
the
others,
or
of
a
biochemical
signal
passing
from
one
terminal
to
the
others
to
induce
the
withdrawal
of
synapses.
In
either
case
this
type
of
hypothesis
suffers
from
the
same
disadvantage
as
the
myelination
hypothe-
sis,
in
that
it
is
difficult
to
explain
direct
and
specific
interactions
between
terminals
separated
by
hundreds
of
microns.
Interactions
directly
involving
the
muscle
fibre.
In
order
to
account
for
the
elimination
of
distributed
inputs
in
seems
necessary
to
invoke
some
type
of
signal
passing
from
the
muscle
fibre
to
the
synapses
upon
it.
It
is
not
clear
what
the
nature
of
the
communication
between
muscle
and
nerve
could
be.
There
might,
for
example,
be
a
signal
from
the
muscle
that
actively
induced
the
removal
of
unwanted
terminals.
A
different
type
of
explanation
is
that
synapses
might
compete
for
some
substance,
provided
in
limited
amounts
by
the
muscle
fibre
and
needed
for
the
survival
of
the
synapse.
The
persistence
of
distant
multiple
synapses
might
then
reflect
the
limits
over
which
synapses
could
compete
for
this
hypothetical
substance.
It
is
evident
that
we
are
now
at
the
stage
of
understanding
only
a
few
of
the
rules
of
the
process
whereby
neuromuscular
connexions
reach
their
mature
state.
These
rules
serve
to
restrict
the
types
of
mechanisms
that
could
control
the
elimination
of
synapses
but
as
yet
they
provide
only
indirect
hints
concerning
the
specific
cells
and
intercellular
signals
in-
volved.
A
major
impetus
behind
the
elimination
process
seems
to
be
a
natural
inability
of
the
motor
neurone
to
maintain
more
than
a
fraction
of
its
original
complement
of
synapses.
The
choice
of
which
synapses
are
to
be
removed
results
from
a
competitive
interaction
between
all
of
the
synapses
within
a
limited
distance
on
each
muscle
fibre.
The
nature
of
the
competition
is
still
not
known
but
it
is
probably
geared
to
favour
the
survival
of
terminals
from
smaller
motor
units
and
it
appears
to
involve
a
signal
from
muscle
to
nerve.

ELIMINATION
OF
SYNAPSES
We
thank
Mr
HAvard
T0nnesen
for
invaluable
technical
assistance.
Drs
Eric
Frank,
Terje
Lomo
and
Wesley
Thompson
have
all
given
good
advice
at
various
stages.
D.V.E.
was
supported
by
a
Helen
Hay
Whitney
fellowship.
M.C.B.
was
European
exchange
fellow
of
the
Royal
Society.
REFERENCES
AKERT,
K.
&
SANDRi,
C.
(1968).
An
electron-microscopic
study
of
zine
iodide-
osmium
impregnation
of
neurons.
I.
Staining
of
synaptic
vesicles
at
cholinergic
junctions.
Brain
Re8.
7,
286-295.
BAGUST,
J.,
LEWIs,
D.
M.
&
WEsTERwAN,
R.
A.
(1973).
Polyneuronal
innervation
of
kitten
skeletal
muscle.
J.
Physiol
229,
241-255.
BENNETr,
M.
R.
&
PErioGRiw,
A.
G.
(1974a).
The
formation
of
synapses
in
striated
muscle
during
development.
J.
Phygiol.
241,
515-545.
BENNETT,
M.
R.
&
PE=IGREW,
A.
G.
(1974b).
The
formation
of
synapses
in
re-
innervated
and
cross-innervated
striated
muscle
during
development.
J.
Phy8iol.
241,
547-573.
BENOrT,
P.
&
CHANGEux,
J.-P.
(1975).
Consequences
of
tenotomy
on
the
evolution
of
multiinnervation
in
developing
rat
soleus
muscle.
Brain
Re8.
99,
345-358.
BLAcKsHAw,
S.
W.
&
WARNER,
A.
E.
(1976).
Low
resistance
junctions
between
mesoderm
cells
during
development
of
trunk
muscles.
J.
Phy8iol.
255,
209-230.
BROWN,
M.
C.,
JANSEN,
J.
K.
S.
&
VAN
EssEN,
D.
(1975).
A
large-scale
reduction
in
motor
neurone
peripheral
fields
during
post-natal
development
in
the
rat.
Acta
phy8iol.
8cand.
95,
3-4A.
BROWN,
M.
C.,
JANSEN,
J.
K.
S.
&
VAN
ESSEN,
D.
(1976).
Polyneuronal
cross-
innervation
of
the
immature
rat
soleus
muscle.
Acta
phygiol.
8cand.
96,
20A.
CHAKuLAs,
J.
J.
&
PAULY,
J.
E.
(1965).
A
study
of
postnatal
growth
of
skeletal
muscle
in
the
rat.
Anat.
Rec.
152,
55-61.
CLOSE,
R.
(1964).
Dynamic
properties
of
fast
and
slow
skeletal
muscles
of
the
rat
during
development.
J.
Phy&iol.
173,
74-95.
CLOSE,
R.
(1967).
Properties
of
motor
units
in
fast
and
slow
skeletal
muscles
of
the
rat.
J.
Phy8iol.
193,
45-55.
DENNIs,
M.
(1975).
Physiological
properties
of
junctions
between
nerve
and
muscle
developing during
salamander
limb
regeneration.
J.
Physiol.
244,
683-702.
EDDS,
M.
V.,
JR
(1953).
Collateral
nerve
regeneration.
Q.
Rev.
Biol.
28,
260-276.
ELSBERG,
C.
A.
(1917).
Experiments
on motor
nerve
regeneration
and
the
direct
neurotization
of
paralyzed
muscles
by
their
own
and
by
foreign
nerves.
Science,
N.Y.
45,
318-320.
FRANK,
E.,
GAUTVIK,
K.
&
SOMMERScHILD,
H.
(1976).
Persistence
of
junctional
acetylcholine
receptors
at
mammalian
neuromuscular
synapses
following
dener-
vation.
Cold
Spring
Harb.
Symp.
quant.
Biol.
40,
275-281.
FiLNK,
E.,
JANSEN,
J.
K.
S.,
L0Mo,
T.
&
WESTGAARD,
R.
H.
(1975).
The
interaction
between
foreign
and
original
motor
nerves
innervating
the
soleus
muscle
of
rats.
J.
Phy8iol.
247,
725-743.
GORDON,
T.,
PERRY,
R.,
TuFEERY,
A.
R.
&
VRBOVA,
G.
(1974).
Possible
mechanisms
determining
synapse
formation
in
developing
skeletal
muscle
of
the
chick.
Cell
&
Tissue
Re8.
155,
13-25.
GumT,
L.
(1962).
Neuromuscular
function
after
regeneration
of
interrupted
nerve
fibres
into
partially
denervated
muscle.
Expl
Neurol.
6,
129-141.
GumAN,
E.
&
HANITJRKovI,
V.
(1966).
Motor
unit
in
old
age.
Nature,
Lond.
209,
921-922.
HUGHES,
A.
F.
W.
(1968).
A8pects
of
Neural
Ontogeny,
pp.
249.
London:
Lagos
Press.
421

422
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
JANsEN,
J.
K.
S.,
L0Mo,
T.,
NIcOLAYsEN,
K.
&
WEsTGAARn,
R.
H.
(1973).
Hyper-
innervation
of
skeletal
muscle
fibres:
dependence
on
muscle
activity.
Science,
N.Y.
181,
559-561.
JANwsi,
J.
K.
S.
&
VAN
EssEN,
D.
(1975).
Re-innervation
of
rat
skeletal
muscle
in
the
presence
of
a-bungarotoxin.
J.
Phy8iol.
250,
651-667.
KARNovsKY,
M.
(1964).
The
localization
of
cholinesterase
in
rat
cardiac
muscle
by
electron
microscopy.
J.
CeU
Bid.
23,
217-232.
LIimmsssER,
L.
&
PUAR,
G.
(1974).
Synaptic
transmission
and
cell
death
during
normal
ganglionic
development.
J.
Phyoiol.
241,
737-749.
LImBINsKA,
L.
&
ZEimENA,
J.
(1966).
Formation
of
new
sites
of
acetylcholine
esterase
activity
in
denervated
muscles
of
young
rats.
Nature,
Lond.
210,
39-41.
REDFERN,
P.
A.
(1970).
Neuromuscular
transmission
in
new-born
rats.
J.
Phy8sol.
209,
701-709.
EXPIANATION
OF
PLATES
PLATE
1
A,
multiple
preterminal
axons
(arrows)
to
single
end-plates
in
an
8-day-old
rat
diaphragm
stained
with
zinc
iodide-osmium.
The
muscle
fibres,
which
run
horizon-
tally
in
the
photograph,
stain
lightly
and
are
not
easily
resolvable.
B,
single
end-plate
stained
for
cholinesterase
on
a
fibre
isolated
from
a
6-day-
old
soleus
muscle.
PLATE
2
A,
cross-section
from
the
middle
of
a
32-day-old
normal
soleus
muscle,
stained
with
Toluidine
blue.
B,
cross-section
of
a
33-day-old
soleus
muscle
partially
denervated
since
day
4.
There
were
321
large
fibres
in
this
muscle,
which
was
innervated
by
only
two motor
axons.
The
scale
applies
to
A
and
B.
C,
multiple'foreign
end-plates
on
a
cross-innervated
muscle
fibre.
The
fibre
was
isolated
from
a
13-day-old
cross-innervated
muscle
after
staining
for
cholinesterase.
Two
end-plates
are
visible
at
a
separation
of
about
150
1m.
D,
double
foreign
end-plates
on an
isolated
fibre
from
a
72-day-old
cross-inner-
vated
muscle
stained
for
cholinesterase.
No
fibres
with
multiple
foreign
e.p.p.
com-
ponents
were
seen
in
this
muscle.

The
Journal
of
Physiology,
Vol.
261,
No.
2
Plate
1
A
~~~~~~~~I
_
50
Um
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
EVEN
(Facing
p.
422)

Uhe
Journal
of
Physiology,
Vol.
261,
No.
2
Plate
2
A
.~~~~~~~nw
__
200
pm
C
50
,m
100pam
M.
C.
BROWN,
J.
K.
S.
JANSEN
AND
D.
VAN
ESSEN
